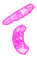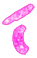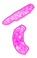Penyakit Puru Akar pada Tanaman Tomat
Tomat (Lycopersicum esculentum Mill) termasuk dalam famili Solanaceae, genus Lycopersicum dan merupakan tanaman musim panas yang memerlukan banyak sinar matahari dan kelembaban tanah yang cukup. Tanaman ini diduga berasal dari Amerika Selatan yang digunakan sebagai bahan makanan di Virginia pada awal tahun 1781 dan dipasarkan di New Orleáns pada tahun 1812. Bagi kesehatan tanaman tomat adalah Sangat penting,buahnya banyak mengandung vitamin A,B1,B2 dan terutama vitamin C,juga mengandung protein dan karbohidrat (Bienz,1980).
Sampai saat ini produksi di Indonesia sekitar 5 ton per hektar.hal ini maíz sangat rendah jika dibanding dengan Filipina yang menghasilkan 9 ton per hektar. Dengan demikian peningkatan produksi dan budidaya tanaman tomat masih memerlukan perhatian yang khusus (Rukmana,1994).
Berbagai usaha dilakukan untuk peningkatan produksi tomat,misalnya pengadaan variets baru,perbaikan pola tanam.Dibalik usaha ini masih banyak kendala yang dihadapi dn masih sulit diatasi,antara lain adanya jasad pengganggu atau patogen. Salah satu patogen yang menyerang tanaman tomat ádalaf nematoda parasit seperti nematoda puru akar (Meloidogyne spp.) (Singh,1978).
Serangan Meloidogyne spp. pada akar dapat menurunkan produksi sebanyak 15 – 60 persen,bahkan dapat mencapai 70 persen bila tanaman yang terserang rentan (Prihanto,1989).Percobaan menunjukkan bahwa dengan sekitar 500 – 800 larva Meloidogyne spp. per ilogram tanah dapat menurunkan produksi sebesar 40 persen (sastrahidayat,1985).Serangan nematoda sering berassosiasi dengan organisme lanilla, misalnya cendawan dan bakteri (Brown,1980).
Khususnya di Indonesia nematoda puru akar pada umumnya dikendalikan dengan penggunaan nematisida Furadan 3 G (Mulyadi,1980) akan tetapi dewasa ini berkembang cara pengendalian organisme pengganggu tanaman dengan menggunakan musuh alami dari organisme pengganggu tanaman tersebut. Cara ini dikenal dengan pengedalian secara hayati. Berkembangna pengendalian secara hayati ini hádala dalam mencari alternatif dari pengendalian secara nimia yang acapkali mengakibatkan dampak negatif bagi kelestarian lingkungan dan kesehatan masyarakat. Keuntungan dari pengendalian secara hayati antara lain bahwa organisme antagonis jarang menyebabkan perubahan ras dari organisme yang dituju sebagaimana sering terjadi kalau menggunakan pestisida. selain hal tersebut aplikasi organismo antagonis tidak sesering aplikasi pesticida,sebab sekali diberikan maka selanjutnya organismo itu sendiri akan terus aktif selama organisme yang dituju tetap ada (James,1985).
Menurut Sarbini (1993) bahwa beberapa cendawan memperlihatkan prospek dalam mengendalikan nematoda Meloidogynei spp. seperti spesies-spesies dari kelas basidiomycetes dan kelas Deutromycetes antara lain genus Gliocladium dan Paecilomyces.
Pengendalian dengan menggunakan Minoriza vesikular arbuskular juga merupakan salah satu pengendalian secara hayati yang dikembangkan saat ini. Penelitian yang telah dilakukan oleh Bagyaray (1975) ternyata Glomus fasciculatus dapat menekan ukuran dan jumlah puru yang ditimbulkan oleh M.incognita dan M.javanica pada tanaman tomat.
Berdasarkan hal tersebut maka dilakukanlah statu penelitian untuk mengetahui kemampuan dari mikoriza dan beberapa cendawan rhizosfer untuk mengendalikan penyakit puru akar yang disebabkan oleh nematoda Meloidogyne spp.
Gejala serangan nematoda pada tanaman menujukkan gejala yang khas pada bagian akar dibawah permukaan tanah. Tumbuhan yang terserang biasanya menunjukkan gejala pertumbuhan ang tidak sehat seperti kerdil dan cenderung mudah layu pada hari-hari panas. Sedangkan akarnya akan mengalami pembengkakan dengan berbagai macam bentuk (Sastrahidayat,1990).
Menurut Bird (1972) akibat serangan nematodo puru akar fungsi alamiah sel tanamn akan terganggu. Sebagai contoh menurunnya laju fotosíntesis, pertumbuhan dan juga produksi. Hal ini diduga bahwa nematoda ini mempengaruhi fisiologi tanaman dengan mengganggu síntesis dan translokasi hormon pertumbuhan yang diproduksi oleh akar.
Apabila sistem perakaran terserang respirasi pada tanaman menigkat,absorbsi oksigen lebih cepat atau seimbang dengan tumbuhan yang sehat serta jaringan xilem menjadi kerdil,sehingga pengambilan air dan nutrisi dari dalam tanah terhambat,akibatnya terjadi kekurangan unsur hara maupun air. Gejala akan tampak pada daun atau bagian lain dari pada tumbuhan (Bird,1972).
Mekanisme hubungan dimulai dari masuknya nematoda ke dalam akar tumbuhan melalui bagian - bagian epidermis yang terletak dekat tudung akar. Nematoda ini mengeluarkan enzim yang dapat menguraikan dinding sel tumbuhan terutama terdiri dari protein,polisakarida seperti pektin,selulase dan hemiselulose serta pati,sukrose dan glicosid menjadi bahan-bahan lain Meloidogyne spp. mengeluarkan enzim selulose yang dapat menghidrolisa selulose enzim endopektin metil transeliminase yang dapat menguraikan pektin.
Dengan terurainya bahan – bahan penyusun dinding sel ini maka dinding sel akan rusak dan terjadilah luka. Selanjutnya nematoda ini bergerak diantara sel-sel atau menembus sel-sel menuju jaringan sel yang terdapat cukup cairan makanan, kemudian menetap dan berkembangbiak kemudian nematoda tersebut masih mengeluarkan enzim proteolitik dengan melepaskan IAA ( Asam Indol Asetat ) yang merupakan heteroauksin tritopan yang diduga membantu terbentuknya puru (Bird,1972).
Mekanisme hubungan dimulai dari masuknya nematoda ke dalam akar tumbuhan melalui bagian - bagian epidermis yang terletak dekat tudung akar. Nematoda ini mengeluarkan enzim yang dapat menguraikan dinding sel tumbuhan terutama terdiri dari protein,polisakarida seperti pektin,selulase dan hemiselulose serta pati, sukrose dan glicosid menjadi bahan-bahan lain Meloidogyne spp. mengeluarkan enzim selulose yang dapat menghidrolisa selulose enzim endopektin metil transeliminase yang dapat menguraikan pektin. Dengan terurainya bahan – bahan penyusun dinding sel ini maka dinding sel akan rusak dan terjadilah luka. Selanjutnya nematoda ini bergerak diantara sel-sel atau menembus sel-sel menuju jaringan sel yang terdapat cukup cairan makanan, kemudian menetap dan berkembangbiak kemudian nematoda tersebut masih mengeluarkan enzim proteolitik dengan melepaskan IAA ( Asam Indol Asetat ) yang merupakan heteroauksin tritopan yang diduga membantu terbentuknya puru (Bird,1972).
Enzim dan hormon yang dikeluarkan oleh nematoda tersebut secara langsung dapat merangsang perkembangan sel. Serangan pertama hanyalah menghentikan diferensiasi sel,sedangkan pada akhirnya serangan sel mulai membesar dan pembagian inti sel terjadi lambat laun dinding sel hilang dan isi protoplasma bersatu membentuk sel besar.Pada akhirnya sel-sel korteks sekeliling sel yang besar mengadakan proliferasi secara cepat untuk menghasilkan puru.Terjadinya puru bukanlah satu-satunya gejala kerusakan oleh nematoda ini. selain hal itu nematoda juga menyebabkan pertukaran bahan-bahan kimia dalam tubuh tumbuhan,misalnya protoplasma sel raksasa mengandung lemak, asam ribonukleat dan protein yang jumlahnya lebih banyak apabila dibandingkan pada sel tumbuhan sehat dan asam cianida (HCN) yang dilepaskan nematoda parasit sangat melemahkan jaringan tumbuhan,sehingga akan menguntungkan bagi penyerangannya patogen lain (Sastrahidayat,1990).
Sastrahidayat (1990) puru akar yang terbentuk tergantung pada spesies tanaman. Pada tanaman sukulen seperti tomat puru berbentuk bulat yang ukurannya dapat mencapai diameter 1,2 cm atau lebih dan kebanyakan terjadi pada akar tunggang tanaman. Selanjutnya Walker (1976) menyatakan bahwa beberapa spesies nematoda puru akar akan membentuk tipe puru akar yang sama,sehingga sulit dibedakan antara satu dengan yang lainnya.
Serangan nematoda menyebabkan pertumbuhan tanaman terhambat,akibatnya produksi dan mutu menurun dan secara ekonomis tidak dapat memberikan hasil yang maksimal jika tidak dilakukan pengendalian yang sesuai (Clarc,1974).
Pengendalian nematoda parasit tanaman dapat dilakukan dengan cara kimia, cara bercocok tanam,pergiliran tanaman, sanitasi dan pengendalian hayati (Mulyadi,1989).pengedalian secara hayati adalah salah satu alternatif sebagai pengganti cara kimia dan cara ini sudah lama dicoba (Horst,1974). Keistimewaan pengendalian hayati adalah terutama mengurangi dampak negatif dari penggunaan pestisida (Mulyadi,1989).
Musuh alami nematoda puru akar sudah banyak diketahui,misalnya didataran tinggi telah ditemukan cendawan Paecilomycetes bilacinus yang menginfeksi telur nematoda puru akar pada tanaman kentang (Anonim,1984).Bacillus penetrans adalah suatu parasit yang dikenal bertahun-tahun berassosiasi dengan Meloidogyne spp. serta beberapa spesies jamur yang menyerang nematoda tanah di Inggris (Kerry,1978). Hasil penelitian sebelumnya menunjukkan bahwa perlakuan dengan cendawan terhadap Meloidogyne spp. dapat menekan jumlah populasi dan intensitas serangan yang memperlihatkan hasil yang baik (Sarbini, 1992).Sarbini (1993) mengemukakan bahwa cendawan parasit telur Meloidogyne spp. terutama dari spesies Gliocladium sp. dan Paecilomyces sp. mempunyai prospek untuk dikembangkan sebagai agen pengendali secara hayati untuk mengendalikan Meloidogyne spp. Hasanuddin (1994) mengemukakan bahwa cendawan rhizosphere antara lain Fusarium sp. dan Trichoderma sp. dapat digunakan untuk mengendalikan nematoda puru akar.
Rabu, 11 April 2012
Rabu, 04 April 2012
cendawan entomopatogen beauveria bassiana
I. PENDAHULUAN
Agens Pengendali Hayati adalah musuh alami berupa Parasitoid, Predator, Entomopatogen dan Agens Antagonis yang telah diketahui mempunyai potensi yang besar sebagai salah satu sarana pengendalian OPT.
Entomopatogen atau penyakit pada serangga hama yang mencakup semua mikroba yaitu Cendawan, bakteri, virus, dan nematoda yang memiliki kemampuan untuk menjadi parasit (pathogen) pada serangga sehingga dapat mematikan serangga yang diinfeksinya. Cendawan adalah mikroba yang mempunyai bentuk tubuh menyerupai benang yang disebut hifa, nampak seperti tumbuhan namun tidak memiliki klorofil.
Mikroba dari jenis jamur adalah Beauveria bassiana, Metarrhizium anisopliae Metarrhizium ftavoviridae Verticillium lecanii dll. dapat mengendalikan Uret, Kepinding tanah, Belalang, Lalat buah dan serangga lainnya.
Kesulitan dan permasalahan utama dalam penerapan dan pengembangan pengendalian hayati adalah modal investasi permulaan yang besar yang harus dikeluarkan untuk kegiatan eksplorasi, penelitian, pengujian dan evaluasi terutama yang menyangkut berbagai aspek dasar baik untuk hama, musuh alami maupun tanaman. Aspek dasar dapat meliputi taksonomi, ekologi, biologi, siklus hidup, dinamika populasi, genetika, fisiologi, dll. Identifikasi yang tepat baik untuk jenis hama maupun musuh alaminya merupakan langkah permulaan yang sangat penting.
Kecuali diperlukan modal, fasilitas yang lengkap juga diperlukan sumber daya manusia terutama para peneliti yang berkualitas dan berpendidikan khusus dan berdedikasi tinggi sesuai dengan yang diperlukan untuk pengembangan teknologi pengendalian hayati. Meskipun ada beberapa ahli yang berpendapat bahwa untuk pengendalian hayati yang penting adalah adanya tenaga peneliti yang berpengalaman dan berdedikasi tinggi serta cukup memiliki rasa seni dan intuisi, namun bagaimanapun untuk keberhasilan pengendalian hayati dalam kerangka PHT diperlukan juga dasar pengetahuan dan teknologi yang mantap.
II. PEMBAHASAN
Adanya kekhawatiran akan pengaruh negatif tentang pemakaian agrokhemikal telah meningkatkan perhatian masyarakat kepada bioinsektisida sebagai alternative teknologi untuk menurunkan populasi hama. Salah satu alternatif pengendalian adalah pemanfaatan jamur penyebab penyakit pada serangga (bioinsectisida), yaitu jamur patogen serangga Beauveria bassiana.
Di Indonesia, hasil-hasil penelitian B. bassiana juga telah banyak dipublikasikan, terutama dari tanaman pangan untuk mengendalikan serangga hama kedelai (Riptortus linearis dan Spodoptera litura), walang sangit pada padi (Leptocoriza acuta) (Prayogo, 2006), Plutella xylostella pada sayursayuran (Hardiyanti, 2006), hama bubuk buah kopi Helopeltis antoni, dan penggerek buah kakaoHypothenemus hampei (Sudarmadji dan Prayogo, dalam Prayogo, 2006).
Saat ini produk bioinsektisida berbahan aktif B. bassiana telah tersedia secara komersial di Indonesia. Meskipun demikian, tampaknya pemanfaatannya di lapang khususnya untuk tanaman perkebunan belum optimal. Padahal,lingkungan mikro tanaman perkebunan sangat ideal bagi perkembangan epizootik cendawan-cendawan entomopatogen, termasuk B. bassiana. Keberlangsungan epizootik cendawan sangat dipengaruhi oleh suhu dan kelembaban lingkungan, dan kriteria ini dapat ditemukan pada tanaman-tanaman perkebunan yang banyak diusahakan di Indonesia. Disamping itu, pemanfaatan cendawan ini dan patogen serangga secara umum dalam pengendalian hama berpotensi memberi keuntungan ekologis jangka panjang terhadap keseimbangan hayati maupun keberlanjutan sistem pertanian.
Pengendalian penyakit tanaman dengan menggunakan agens pengendali hayati muncul karena kekhawatiran masyarakat dunia akibat penggunaan pestisida kimia sintetis. Adanya kekhawatiran tersebut membuat pengendalian hayati menjadi salah satu pilihan cara mengendalikan patogen tanaman yang harus dipertimbangkan.
Agensia hayati adalah setiap organisme yang meliputi spesies, subspesies, varietas, semua jenis serangga, nematoda, protozoa, cendawan (fungi), bakteri, virus, mikoplasma, serta organisme lainnya dalam semua tahap perkembangannya yang dapat dipergunakan untuk keperluan pengendalian hama dan penyakit atau organisme pengganggu, proses produksi, pengolahan hasil pertanian, dan berbagai keperluan lainnya (Permentan No. 411 tahun 1995).
Beauvaria bassiana merupakan cendawan entomopatogen yaitu cendawan yang dapat menimbulkan penyakit pada serangga. Beauveria bassiana secara alami terdapat di dalam tanah sebagai jamur saprofit. Pertumbuhan jamur di dalam tanah sangat dipengaruhi oleh kondisi tanah, seperti kandungan bahan organik, suhu, kelembapan, kebiasaan makan serangga, adanya pestisida sintetis, dan waktu aplikasi. Secara umum, suhu di atas 30 °C, kelembapan tanah yang berkurang dan adanya antifungal atau pestisida dapat menghambat pertumbuhannya.
Beauveria bassiana membunuh hama melalui infeksi sebagai akibat dari serangga yang kontak dengan spora jamur. Serangga dapat kontak dengan spora jamur melalui beberapa cara: semprotan jamur menempel pada tubuh serangga, serangga bergerak pada permukaan tanaman yang sudah terinfeksi jamur, atau dengan memakan jaringan tanaman yang telah diperlakukan dengan jamur. Setelah spora jamur melekat pada kulit serangga (kutikula), mereka berkecambah membentuk struktur (hifa) yang menembus tubuh serangga dan berkembang biak. Proses ini memakan waktu 3-5 hari sampai akhirnya serangga mati, bangkai yang terinfeksi dapat berfungsi sebagai sumber spora untuk penyebaran sekunder jamur. Keberhasilan penyemprotan menggunakan jamur Beauveria bassiana bergantung pada kerentanan spesies yang bersangkutan, tingkat populasi hama, dan kondisi lingkungan pada saat aplikasi, serta sumber daya manusia itu sendiri.
Mekanisme Infeksi Beauveria Bassiana
B. bassiana memproduksi toksin yang disebut beauvericin (Kučera dan Samšiňáková, 1968). Antibiotik ini dapat menyebabkan gangguan pada fungsi hemolimfa dan nukleus serangga, sehingga mengakibatkan pembengkakan yang disertai pengerasan pada serangga yang terinfeksi. Selain secara kontak, B. bassiana juga dapat menginfeksi serangga melalui inokulasi atau kontaminasi pakan. Broome et al. (1976) menyatakan bahwa 37% dari konidia B. bassiana yang dicampurkan ke dalam pakan semut api, Selenopsis richteri, berkecambah di dalam saluran pencernaan inangnya dalam waktu 72 jam, sedangkan hifanya mampu menembus dinding usus antara 60-72 jam. Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat. Serangga yang mati tidak selalu disertai gejala pertumbuhan spora. Contohnya, aphid yang terinfeksi B.bassiana hanya mengalami pembengkakan tanpa terjadi perubahan warna. Demikian pula tempayak lalat yang terinfeksi B. bassiana sering ditemukan secara berkelompok pada ujung-ujung rerumputan (Plate, 1976).
Kematian serangga biasanya disebabkan oleh kerusakan jaringan secara menyeluruh, dan atau karena toksin yang diproduksi oleh cendawan. Menurut Cheung dan Grula (1982), penyakit white muscardine yang menyerang saluran pencernaan Heliothis zea mengakibatkan gangguan nutrisi hingga kematian. Serangga yang terbunuh tubuhnya akan berwarna putih karena ditumbuhi konidia B. bassiana. Jumlah konidia yang dapat dihasilkan oleh satu serangga ditentukan oleh besar kecilnya ukuran serangga tersebut. Setiap serangga terinfeksi B. bassiana akan efektif menjadi sumber infeksi bagi serangga sehat di sekitarnya.
Seperti cendawan lain, perrtumbuhan B. bassiana juga sangat ditentukan oleh kelembapan lingkungan. Namun demikian, cendawan ini juga memiliki fase resisten yang dapat mempertahankan kemampuannya menginfeksi inang pada kondisi kering. Keberadaan epizootiknya di alam menyebabkan B. bassiana secara cepat menginfeksi populasi serangga hingga menyebabkan kematian. Selain itu, kemampuan penetrasinya yang tinggi pada tubuh serangga menyebabkan cendawan ini juga dengan mudah menginfeksi serangga hama pengisap, seperti aphid (Aphis sp.) dan kutu putih Bemisia spp. yang tidak mudah terinfeksi oleh bakteri maupun virus.
Faktor lingkungan, terutama kelembaban dan temperatur serta sedikit cahaya sangat penting perannya dalam proses infeksi dan sporulasi cendawan entomopatogen (Roberts dan Campbell, 1977; McCoy et al., 1988). Temperatur optimum untuk perkembangan, patogenisitas, dan kelulusan hidup cendawan umumnya antara 20-30°C (McCoy et al., 1988). Untuk perkecambahan konidia dan sporulasi pada permukaan tubuh serangga dibutuhkan kelembaban sangat tinggi (> 90% RH), terutama kelembaban di lingkungan mikro sekitar konidia sangat penting perannya dalam proses perkecambahan dan produksi konidia (Millstein et al., 1983; Nordin et al., 1983). Tetapi sebaliknya untuk melepaskan konidia B. bassiana dari konidifor hanya dibutuhkan kelembaban sekitar 50% (Gottwald dan Tedders, 1982).
Meskipun pengaruh cahaya terhadap infeksi cendawan belum diketahui secara jelas, tetapi intensitas sinar ultraviolet tertentu dapat merusak konidia cendawan (Callaghan, 1969). Fuxa (1987) menyatakan bahwa intensitas cahaya matahari dengan rata-rata panjang gelombang antara 290-400 nm cukup efektif menurunkan persistensi deposit konidia pada pertanaman. Sementara Ignoffo et al. (1977) mengemukakan bahwa waktu paruh (half-life) sebagian besar spora cendawan yang terekspos cahaya buatan dengan panjang gelombang mendekati panjang gelombang sinar matahari (290-400 nm) hanya sekitar 1-4 jam, tetapi kenyataannya di lapang waktu paruh dapat mencapai lebih dari 4 jam.
Umumnya cendawan entomopatogen membutuhkan lingkungan yang lembab untuk dapat menginfeksi serangga, oleh karena itu epizootiknya di alam biasanya terbentuk pada saat kondisi lingkungan lembab atau basah. Keefektifan B. bassiana menginfeksi serangga hama tergantung pada spesies atau strain cendawan, dan kepekaan stadia serangga pada tingkat kelembaban lingkungan, struktur tanah (untuk serangga dalam tanah), dan temperatur yang tepat. Selain itu, harus terjadi kontak antara spora B. bassiana yang diterbangkan angin atau terbawa air dengan serangga inang agar terjadi infeksi. Plate (1976) juga menyatakan bahwa epizootik cendawan yang terbentuk secara alami efektif mengendalikan populasi aphid, tempayak lalat yang menyerang perakaran tanaman, belalang, dan thrip, disamping juga potensial sebagai faktor mortalitas utama aphid yang menyerang kentang dan tanaman inang lainnya.
Konidia merupakan unit B. bassiana yang paling infektif dan stabil untuk aplikasi di lapang dibandingkan dengan hifa maupun blastosporanya (Soper dan Ward, 1981; Feng et al., 1994). Konidia yang diaplikasikan dapat berupa suspensi (tidak diformulasi), formulasi butiran, dan bentuk pellet, dan ketiganya memperlihatkan hasil pengendalian yang cukup nyata. Stimac et al. (1993) menyatakan bahwa aplikasi konidia B. bassiana dengan cara sprinkle dan disemprotkan pada permukaan tanah sangat efektif menyebabkan mortalitas hama sasaran. Mortalitas hama semut api, Selenopsis invicta, yang dikendalikan dengan B. bassiana tertinggi mulai 3-8 hari setelah perlakuan. Sedangkan enkapsulasi (pellet) konidia B. bassiana dengan menggunakan kalsium alginat juga efektif meningkatkan mortalitas S. invicta (White, 1995), karena enkapsulasi menyebabkan konidia lebih stabil di dalam tanah.
Beberapa senyawa metabolit sekunder diproduksi oleh B. bassiana, seperti beauvericin, bassianin, bassiacridin, bassianolide, beauverolides, tenellin, dan oosporein (Strasser et al., 2000; Vey et al., 2001; Quesada-Moraga dan Vey, 2004). Senyawa metabolit sekunder ini dapat dihasilkan oleh B. bassiana pada epizootik di alam (tanah) maupun pada epizootik buatan (di laboratorium). Meskipun demikian, hingga saat ini belum ada laporan tentang tercemarnya rantai makanan oleh senyawa metabolit sekunder, atau terakumulasi di alam sebagai limbah epizootik B. bassiana (Vey et al., 2001).
Penggunaan B. bassiana dalam pengendalian hama telah diuji secara luas di berbagai negara. Hasil uji toksikologi terhadap salah satu produk B. bassiana, Botanigard, menunjukkan bahwa produk tersebut tidak menimbulkan dampak negatif yang berhubungan dengan patogenisitas dan toksisitasnya, sehingga produk tersebut digunakan secara aman selama lebih dari 10 tahun di Amerika Serikat dan juga di beberapa negara lain (US EPA, 2006).
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna. Namun, Plate (1976) mengungkapkan bahwa tingkat kepekaan serangga bukan sasaran terhadap infeksi B. bassiana sangat ditentukan oleh virulensi dan patogenisitas cendawan, serta spesies serangga inang. Selain itu, perbedaan fisiologis dan ekologis inang juga mempengaruhi infeksi B. bassiana. Misalnya, serangga bukan sasaran yang mudah terinfeksi B. bassiana di laboratorium tidak akan serta merta terinfeksi pada kondisi lapang. Ludwig dan Oetting (2001) menegaskan bahwa beberapa serangga musuh alami yang peka terhadap infeksi B. bassiana di laboratorium ternyata mengalami infeksi sangat rendah pada uji di rumah kaca. Disamping itu, hasil uji ekotoksikologi terhadap produk Botanigard menunjukkan bahwa risiko secara ekologis yang diperlihatkan oleh serangga bukan sasaran yang diperlakukan dengan formulasi B. bassiana sangat rendah (US EPA, 2006).
Dihubungkan dengan keamanan secara hayati, cendawan entomopatogen dikelompokkan menjadi cendawan dengan kisaran inang spesifik dan yang kisaran inangnya luas (MacLeod, 1963). Cendawan yang memiliki kisaran inang spesifik umumnya menjadi parasit sejati (obligat) dan bersifat sangat virulen tarhadap inang. Sebaliknya yang kisaran inangnya luas sebagian besar merupakan patogen fakultatif, bersifat saprofit, dan cenderung kurang patogenik (Goettel et al., 1990), dan biasanya virulensinya tinggi hanya pada spesies inang dari mana cendawan tersebut pertama kali diisolasi. Contoh, B. bassiana yang diisolasi dari ulat H. armigera akan lebih patogenik pada inangnya tersebut dibanding dengan inang-inangnya yang lain. Selain itu, cendawan yang kisaran inangnya lebih luas justru menjadi lebih spesifik menginfeksi inang jika di lapang. Hal ini dapat terjadi kemungkinan karena dipengaruhi oleh interaksi antara faktor abiotik dan biotik di lapang, sehingga serangga yang mudah terinfeksi di laboratorium belum tentu mudah juga terinfeksi di lapang. Oleh karena itu, kemungkinan terinfeksinya serangga bukan sasaran oleh B. bassiana di lapang sangat kecil. Dengan demikian, aplikasi B. bassiana di lapang cenderung aman bagi musuh alami atau serangga berguna lainnya. Infeksi B. bassiana pada manusia sangat jarang terjadi. Meskipun demikian, dilaporkan ada dua kasus infeksi B. bassiana yang menyebabkan mikosis pada manusia (Henke et al., 2002; Tucker et al., 2004). Namun infeksi tersebut terjadi pada kondisi kesehatan manusia yang sangat buruk akibat penyakit leukimia akut.
Pengujian Botanigard terhadap mamalia, burung, dan ikan juga tidak menunjukkan pengaruh negatif terhadap perkembangan hewan-hewan tersebut (US EPA, 2006). Demikian pula pengujian terhadap sejumlah reptil maupun vertebrata yang membuktikan bahwa B. bassiana tidak menginfeksi keduanya (Georg et al., 1962; Fromtling et al., 1979; Gonzalez et al., 1995).
Cukup banyak tersedia bahan untuk media alami perbanyakan B. bassiana, antara lain: beras, gandum, kedelai, jagung, padi-padian, sorghum, kentang, roti, dan kacang-kacangan. Bahan mana yang akan digunakan tergantung pada beberapa faktor, termasuk kemudahan memperoleh bahan tersebut, biaya, dan strain isolat yang akan diperbanyak. Dalam perbanyakan B. bassiana dengan bahan-bahan alami, untuk menghasilkan konidia dalam jumlah maksimal diperlukan media dengan partikel yang permukaannya lebih luas. Bahan media yang cenderung menggumpal akan memiliki luas permukaan yang sempit, sehingga produksi konidia juga sedikit. Media yang ideal adalah media yang tidak hanya mempunyai partikel dengan permukaan luas, tetapi juga yang dapat mempertahankan keutuhan partikel selama proses produksi (Maheva et al., 1984; Bradley et al., 1992).
Tiga jenis bahan media alami yang telah dicoba dalam perbanyakan B. bassiana skala besar di New Zealand adalah beras, gandum, dan barley. Hasilnya, beras merupakan media paling sesuai bagi perkembangan B. bassiana dengan produktivitas konidia tertinggi mencapai 4,38 x 109 konidia/g beras (Nelson dan Glare, 1996). Penggunaan berbagai jenis sereal, selain beras, sebagai media perbanyakan B. bassiana perlu dipertimbangkan mengingat kandungan nutrisinya yang sangat bervariasi (Jenkins et al., 1998). Perbedaan kandungan nutrisi ini sangat mempengaruhi produksi konidia, terutama per kelompok produksi (batch). Oleh karena itu, pemilihan bahan media perbanyakan harus dilakukan secara tepat, terutama memilih bahan yang memiliki kemampuan produksi konidia secara konsisten dalam kelompok-kelompok produksi. Hasil penelitian lain juga membuktikan bahwa beras putih merupakan bahan media perbanyakan B. bassiana yang tepat karena produksi konidia yang tinggi (Alves dan Pereira, 1989; Mendonca, 1992; Ibrahim dan Low, 1993; Milner et al., 1993). Hal tersebut
Menunjukkan bahwa kombinasi faktor-faktor produkai sangat kompatibel, termasuk keseimbangan nutrisi dalam bahan media, biaya produksi, kemudahan memperoleh bahan, karakter fisik bahan, seperti ukuran, bentuk, dan keutuhan bahan baik sebelum maupun setelah pengkolonisasian konidia.
Dalam perbanyakan, temperatur inkubasi dan cahaya sangat menentukan produktivitas konidia. Temperatur optimal setiap cendawan bervariasi tidak saja antar spesies, tetapi juga antar isolat (Thomas dan Jenkins, 1997; Alasoadura, 1963). Temperatur optimal untuk perkecambahan konidia adalah 25-30°C, dengan temperatur minimum 10°C dan maksimum 32°C. Sedangkan pH optimal untuk pertumbuhan adalah 5,7-5,9, tapi idealnya pH 7-8 (Goral dan Lappa, 1972). Beberapa cendawan membutuhkan cahaya untuk proses sporulasi, sedangkan cendawan lainnya tidak terpengaruh oleh cahaya. Tetapi ada pula cendawan yang sporulasinya terhambat pada tingkat intensitas cahaya tertentu (Vouk dan Klas, 1931). Penelitian terdahulu membuktikan bahwa B. bassiana yang diproduksi di lingkungan tanpa cahaya (gelap) konidianya cenderung berukuran lebih besar dan lebih virulen dibanding yang diproduksi pada tempat terang (Humphreys et al., 1989; Williams, 1959). Hal ini penting sebagai bahan pertimbangan dalam memilih kemasan yang sesuai apabila biakan cendawan harus dibawa ke luar areal perbanyakan. Selain itu yang lebih penting dalam perbanyakan B. bassiana untuk skala komersial adalah kesesuaian produk dengan teknik formulasi dan aplikasinya.
Umumnya produk B. bassiana diformulasi dalam bentuk bubuk (powder) dan merupakan formulasi paling efektif memicu kontak dengan hama sasaran (Stimac et al., 1993). Formulasi B. bassiana berupa pellet hasil enkapsulasi miselium selain efektif untuk meningkatkan mortalitas hama juga untuk mengurangi kompetisi dengan mikroba lain, sehingga meningkatkan daya hidup B. bassiana (White, 1995).
Dalam jurnal diketahui kalau penambahan tepung tapioka sebanyak 1 g dan suhu penyimpanan 5OC, mempunyai potensi yang baik dalam mempertahankan viabilitas spora B. bassiana sekurang-kurangnya sampai dua bulan penyimpanan. Hal tersebut disebabkan oleh kemampuan daya simpan tepung tapioka yang lebih baik daripada tepung beras dan maizena serta didukung oleh suhu rendah yang sesuai untuk mempertahankan viabilitas spora B. bassiana. Terjadi penurunan viabilitas spora kering B. bassiana lebih cepat, seiring dengan semakin meningkatnya dosis pembawa, suhu dan lama penyimpanan. Penyimpanan pada suhu 5OC menunjukkan kemampuan mempertahankan viabilitas spora B. bassiana kering murni lebih lama daripada kondisi suhu 23C dan suhu 29C.
II. PENUTUP
Kesimpulan
Dari pembahasan di atas kita dapat menyimpulkan bahwa:
Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat
Ada beberapa faktor yang mempengaruhi mekanisme infeksi cendawan entomopatogen Beauveria bassiana, faktor yang meliputi diantaranya kelembaban, suhu, iklim, dan tingkat kekebalan tubuh dari inang
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna.
DAFTAR PUSTAKA
Anonim, 2012. Perbanyakan Cendawan Entomopatogen. http: www. Pengendalian%agents-hayati.com.html.diakses pada tanggal 21 maret 2012
Anonim, 2012. Prospek Beauveria bassiana sebagai bioinsectisida. http//:www. Miss Biology Prospek Beauveria bassiana sebagai bioinsectisida.htm. diakses pada tanggal 21 maret 2012
Anonim, 2012. Beauveria bassiana . Wikipedia bahasa Indonesia, ensiklopedia bebas.html. diakses pada tanggal 21 maret 2012
Balai Penelitian Tanaman Hias. 2006. Beauveria bassiana pengendali hama tanaman. Warta Penelitian dan Pengembangan Pertanian Indonesia 28(1): 11-12.
EOL. 2000. Beauveria bassiana Bals. -Criv. Vuill. 1912.[terhubung berkala] http://www.eol.org/pages/160292 . diakses pada tanggal 21 maret 2012
McCoy C, Quintela ED, Faria M de. 1990. Environmental persistence of entomopathogenic fungi. [terhubung berkala]. http://www.lsuagcenter.com/s265/mccoy.htm. diakses pada tanggal 21 maret 2012
I. PENDAHULUAN
Agens Pengendali Hayati adalah musuh alami berupa Parasitoid, Predator, Entomopatogen dan Agens Antagonis yang telah diketahui mempunyai potensi yang besar sebagai salah satu sarana pengendalian OPT.
Entomopatogen atau penyakit pada serangga hama yang mencakup semua mikroba yaitu Cendawan, bakteri, virus, dan nematoda yang memiliki kemampuan untuk menjadi parasit (pathogen) pada serangga sehingga dapat mematikan serangga yang diinfeksinya. Cendawan adalah mikroba yang mempunyai bentuk tubuh menyerupai benang yang disebut hifa, nampak seperti tumbuhan namun tidak memiliki klorofil.
Mikroba dari jenis jamur adalah Beauveria bassiana, Metarrhizium anisopliae Metarrhizium ftavoviridae Verticillium lecanii dll. dapat mengendalikan Uret, Kepinding tanah, Belalang, Lalat buah dan serangga lainnya.
Kesulitan dan permasalahan utama dalam penerapan dan pengembangan pengendalian hayati adalah modal investasi permulaan yang besar yang harus dikeluarkan untuk kegiatan eksplorasi, penelitian, pengujian dan evaluasi terutama yang menyangkut berbagai aspek dasar baik untuk hama, musuh alami maupun tanaman. Aspek dasar dapat meliputi taksonomi, ekologi, biologi, siklus hidup, dinamika populasi, genetika, fisiologi, dll. Identifikasi yang tepat baik untuk jenis hama maupun musuh alaminya merupakan langkah permulaan yang sangat penting.
Kecuali diperlukan modal, fasilitas yang lengkap juga diperlukan sumber daya manusia terutama para peneliti yang berkualitas dan berpendidikan khusus dan berdedikasi tinggi sesuai dengan yang diperlukan untuk pengembangan teknologi pengendalian hayati. Meskipun ada beberapa ahli yang berpendapat bahwa untuk pengendalian hayati yang penting adalah adanya tenaga peneliti yang berpengalaman dan berdedikasi tinggi serta cukup memiliki rasa seni dan intuisi, namun bagaimanapun untuk keberhasilan pengendalian hayati dalam kerangka PHT diperlukan juga dasar pengetahuan dan teknologi yang mantap.
II. PEMBAHASAN
Adanya kekhawatiran akan pengaruh negatif tentang pemakaian agrokhemikal telah meningkatkan perhatian masyarakat kepada bioinsektisida sebagai alternative teknologi untuk menurunkan populasi hama. Salah satu alternatif pengendalian adalah pemanfaatan jamur penyebab penyakit pada serangga (bioinsectisida), yaitu jamur patogen serangga Beauveria bassiana.
Di Indonesia, hasil-hasil penelitian B. bassiana juga telah banyak dipublikasikan, terutama dari tanaman pangan untuk mengendalikan serangga hama kedelai (Riptortus linearis dan Spodoptera litura), walang sangit pada padi (Leptocoriza acuta) (Prayogo, 2006), Plutella xylostella pada sayursayuran (Hardiyanti, 2006), hama bubuk buah kopi Helopeltis antoni, dan penggerek buah kakaoHypothenemus hampei (Sudarmadji dan Prayogo, dalam Prayogo, 2006).
Saat ini produk bioinsektisida berbahan aktif B. bassiana telah tersedia secara komersial di Indonesia. Meskipun demikian, tampaknya pemanfaatannya di lapang khususnya untuk tanaman perkebunan belum optimal. Padahal,lingkungan mikro tanaman perkebunan sangat ideal bagi perkembangan epizootik cendawan-cendawan entomopatogen, termasuk B. bassiana. Keberlangsungan epizootik cendawan sangat dipengaruhi oleh suhu dan kelembaban lingkungan, dan kriteria ini dapat ditemukan pada tanaman-tanaman perkebunan yang banyak diusahakan di Indonesia. Disamping itu, pemanfaatan cendawan ini dan patogen serangga secara umum dalam pengendalian hama berpotensi memberi keuntungan ekologis jangka panjang terhadap keseimbangan hayati maupun keberlanjutan sistem pertanian.
Pengendalian penyakit tanaman dengan menggunakan agens pengendali hayati muncul karena kekhawatiran masyarakat dunia akibat penggunaan pestisida kimia sintetis. Adanya kekhawatiran tersebut membuat pengendalian hayati menjadi salah satu pilihan cara mengendalikan patogen tanaman yang harus dipertimbangkan.
Agensia hayati adalah setiap organisme yang meliputi spesies, subspesies, varietas, semua jenis serangga, nematoda, protozoa, cendawan (fungi), bakteri, virus, mikoplasma, serta organisme lainnya dalam semua tahap perkembangannya yang dapat dipergunakan untuk keperluan pengendalian hama dan penyakit atau organisme pengganggu, proses produksi, pengolahan hasil pertanian, dan berbagai keperluan lainnya (Permentan No. 411 tahun 1995).
Beauvaria bassiana merupakan cendawan entomopatogen yaitu cendawan yang dapat menimbulkan penyakit pada serangga. Beauveria bassiana secara alami terdapat di dalam tanah sebagai jamur saprofit. Pertumbuhan jamur di dalam tanah sangat dipengaruhi oleh kondisi tanah, seperti kandungan bahan organik, suhu, kelembapan, kebiasaan makan serangga, adanya pestisida sintetis, dan waktu aplikasi. Secara umum, suhu di atas 30 °C, kelembapan tanah yang berkurang dan adanya antifungal atau pestisida dapat menghambat pertumbuhannya.
Beauveria bassiana membunuh hama melalui infeksi sebagai akibat dari serangga yang kontak dengan spora jamur. Serangga dapat kontak dengan spora jamur melalui beberapa cara: semprotan jamur menempel pada tubuh serangga, serangga bergerak pada permukaan tanaman yang sudah terinfeksi jamur, atau dengan memakan jaringan tanaman yang telah diperlakukan dengan jamur. Setelah spora jamur melekat pada kulit serangga (kutikula), mereka berkecambah membentuk struktur (hifa) yang menembus tubuh serangga dan berkembang biak. Proses ini memakan waktu 3-5 hari sampai akhirnya serangga mati, bangkai yang terinfeksi dapat berfungsi sebagai sumber spora untuk penyebaran sekunder jamur. Keberhasilan penyemprotan menggunakan jamur Beauveria bassiana bergantung pada kerentanan spesies yang bersangkutan, tingkat populasi hama, dan kondisi lingkungan pada saat aplikasi, serta sumber daya manusia itu sendiri.
Mekanisme Infeksi Beauveria Bassiana
B. bassiana memproduksi toksin yang disebut beauvericin (Kučera dan Samšiňáková, 1968). Antibiotik ini dapat menyebabkan gangguan pada fungsi hemolimfa dan nukleus serangga, sehingga mengakibatkan pembengkakan yang disertai pengerasan pada serangga yang terinfeksi. Selain secara kontak, B. bassiana juga dapat menginfeksi serangga melalui inokulasi atau kontaminasi pakan. Broome et al. (1976) menyatakan bahwa 37% dari konidia B. bassiana yang dicampurkan ke dalam pakan semut api, Selenopsis richteri, berkecambah di dalam saluran pencernaan inangnya dalam waktu 72 jam, sedangkan hifanya mampu menembus dinding usus antara 60-72 jam. Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat. Serangga yang mati tidak selalu disertai gejala pertumbuhan spora. Contohnya, aphid yang terinfeksi B.bassiana hanya mengalami pembengkakan tanpa terjadi perubahan warna. Demikian pula tempayak lalat yang terinfeksi B. bassiana sering ditemukan secara berkelompok pada ujung-ujung rerumputan (Plate, 1976).
Kematian serangga biasanya disebabkan oleh kerusakan jaringan secara menyeluruh, dan atau karena toksin yang diproduksi oleh cendawan. Menurut Cheung dan Grula (1982), penyakit white muscardine yang menyerang saluran pencernaan Heliothis zea mengakibatkan gangguan nutrisi hingga kematian. Serangga yang terbunuh tubuhnya akan berwarna putih karena ditumbuhi konidia B. bassiana. Jumlah konidia yang dapat dihasilkan oleh satu serangga ditentukan oleh besar kecilnya ukuran serangga tersebut. Setiap serangga terinfeksi B. bassiana akan efektif menjadi sumber infeksi bagi serangga sehat di sekitarnya.
Seperti cendawan lain, perrtumbuhan B. bassiana juga sangat ditentukan oleh kelembapan lingkungan. Namun demikian, cendawan ini juga memiliki fase resisten yang dapat mempertahankan kemampuannya menginfeksi inang pada kondisi kering. Keberadaan epizootiknya di alam menyebabkan B. bassiana secara cepat menginfeksi populasi serangga hingga menyebabkan kematian. Selain itu, kemampuan penetrasinya yang tinggi pada tubuh serangga menyebabkan cendawan ini juga dengan mudah menginfeksi serangga hama pengisap, seperti aphid (Aphis sp.) dan kutu putih Bemisia spp. yang tidak mudah terinfeksi oleh bakteri maupun virus.
Faktor lingkungan, terutama kelembaban dan temperatur serta sedikit cahaya sangat penting perannya dalam proses infeksi dan sporulasi cendawan entomopatogen (Roberts dan Campbell, 1977; McCoy et al., 1988). Temperatur optimum untuk perkembangan, patogenisitas, dan kelulusan hidup cendawan umumnya antara 20-30°C (McCoy et al., 1988). Untuk perkecambahan konidia dan sporulasi pada permukaan tubuh serangga dibutuhkan kelembaban sangat tinggi (> 90% RH), terutama kelembaban di lingkungan mikro sekitar konidia sangat penting perannya dalam proses perkecambahan dan produksi konidia (Millstein et al., 1983; Nordin et al., 1983). Tetapi sebaliknya untuk melepaskan konidia B. bassiana dari konidifor hanya dibutuhkan kelembaban sekitar 50% (Gottwald dan Tedders, 1982).
Meskipun pengaruh cahaya terhadap infeksi cendawan belum diketahui secara jelas, tetapi intensitas sinar ultraviolet tertentu dapat merusak konidia cendawan (Callaghan, 1969). Fuxa (1987) menyatakan bahwa intensitas cahaya matahari dengan rata-rata panjang gelombang antara 290-400 nm cukup efektif menurunkan persistensi deposit konidia pada pertanaman. Sementara Ignoffo et al. (1977) mengemukakan bahwa waktu paruh (half-life) sebagian besar spora cendawan yang terekspos cahaya buatan dengan panjang gelombang mendekati panjang gelombang sinar matahari (290-400 nm) hanya sekitar 1-4 jam, tetapi kenyataannya di lapang waktu paruh dapat mencapai lebih dari 4 jam.
Umumnya cendawan entomopatogen membutuhkan lingkungan yang lembab untuk dapat menginfeksi serangga, oleh karena itu epizootiknya di alam biasanya terbentuk pada saat kondisi lingkungan lembab atau basah. Keefektifan B. bassiana menginfeksi serangga hama tergantung pada spesies atau strain cendawan, dan kepekaan stadia serangga pada tingkat kelembaban lingkungan, struktur tanah (untuk serangga dalam tanah), dan temperatur yang tepat. Selain itu, harus terjadi kontak antara spora B. bassiana yang diterbangkan angin atau terbawa air dengan serangga inang agar terjadi infeksi. Plate (1976) juga menyatakan bahwa epizootik cendawan yang terbentuk secara alami efektif mengendalikan populasi aphid, tempayak lalat yang menyerang perakaran tanaman, belalang, dan thrip, disamping juga potensial sebagai faktor mortalitas utama aphid yang menyerang kentang dan tanaman inang lainnya.
Konidia merupakan unit B. bassiana yang paling infektif dan stabil untuk aplikasi di lapang dibandingkan dengan hifa maupun blastosporanya (Soper dan Ward, 1981; Feng et al., 1994). Konidia yang diaplikasikan dapat berupa suspensi (tidak diformulasi), formulasi butiran, dan bentuk pellet, dan ketiganya memperlihatkan hasil pengendalian yang cukup nyata. Stimac et al. (1993) menyatakan bahwa aplikasi konidia B. bassiana dengan cara sprinkle dan disemprotkan pada permukaan tanah sangat efektif menyebabkan mortalitas hama sasaran. Mortalitas hama semut api, Selenopsis invicta, yang dikendalikan dengan B. bassiana tertinggi mulai 3-8 hari setelah perlakuan. Sedangkan enkapsulasi (pellet) konidia B. bassiana dengan menggunakan kalsium alginat juga efektif meningkatkan mortalitas S. invicta (White, 1995), karena enkapsulasi menyebabkan konidia lebih stabil di dalam tanah.
Beberapa senyawa metabolit sekunder diproduksi oleh B. bassiana, seperti beauvericin, bassianin, bassiacridin, bassianolide, beauverolides, tenellin, dan oosporein (Strasser et al., 2000; Vey et al., 2001; Quesada-Moraga dan Vey, 2004). Senyawa metabolit sekunder ini dapat dihasilkan oleh B. bassiana pada epizootik di alam (tanah) maupun pada epizootik buatan (di laboratorium). Meskipun demikian, hingga saat ini belum ada laporan tentang tercemarnya rantai makanan oleh senyawa metabolit sekunder, atau terakumulasi di alam sebagai limbah epizootik B. bassiana (Vey et al., 2001).
Penggunaan B. bassiana dalam pengendalian hama telah diuji secara luas di berbagai negara. Hasil uji toksikologi terhadap salah satu produk B. bassiana, Botanigard, menunjukkan bahwa produk tersebut tidak menimbulkan dampak negatif yang berhubungan dengan patogenisitas dan toksisitasnya, sehingga produk tersebut digunakan secara aman selama lebih dari 10 tahun di Amerika Serikat dan juga di beberapa negara lain (US EPA, 2006).
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna. Namun, Plate (1976) mengungkapkan bahwa tingkat kepekaan serangga bukan sasaran terhadap infeksi B. bassiana sangat ditentukan oleh virulensi dan patogenisitas cendawan, serta spesies serangga inang. Selain itu, perbedaan fisiologis dan ekologis inang juga mempengaruhi infeksi B. bassiana. Misalnya, serangga bukan sasaran yang mudah terinfeksi B. bassiana di laboratorium tidak akan serta merta terinfeksi pada kondisi lapang. Ludwig dan Oetting (2001) menegaskan bahwa beberapa serangga musuh alami yang peka terhadap infeksi B. bassiana di laboratorium ternyata mengalami infeksi sangat rendah pada uji di rumah kaca. Disamping itu, hasil uji ekotoksikologi terhadap produk Botanigard menunjukkan bahwa risiko secara ekologis yang diperlihatkan oleh serangga bukan sasaran yang diperlakukan dengan formulasi B. bassiana sangat rendah (US EPA, 2006).
Dihubungkan dengan keamanan secara hayati, cendawan entomopatogen dikelompokkan menjadi cendawan dengan kisaran inang spesifik dan yang kisaran inangnya luas (MacLeod, 1963). Cendawan yang memiliki kisaran inang spesifik umumnya menjadi parasit sejati (obligat) dan bersifat sangat virulen tarhadap inang. Sebaliknya yang kisaran inangnya luas sebagian besar merupakan patogen fakultatif, bersifat saprofit, dan cenderung kurang patogenik (Goettel et al., 1990), dan biasanya virulensinya tinggi hanya pada spesies inang dari mana cendawan tersebut pertama kali diisolasi. Contoh, B. bassiana yang diisolasi dari ulat H. armigera akan lebih patogenik pada inangnya tersebut dibanding dengan inang-inangnya yang lain. Selain itu, cendawan yang kisaran inangnya lebih luas justru menjadi lebih spesifik menginfeksi inang jika di lapang. Hal ini dapat terjadi kemungkinan karena dipengaruhi oleh interaksi antara faktor abiotik dan biotik di lapang, sehingga serangga yang mudah terinfeksi di laboratorium belum tentu mudah juga terinfeksi di lapang. Oleh karena itu, kemungkinan terinfeksinya serangga bukan sasaran oleh B. bassiana di lapang sangat kecil. Dengan demikian, aplikasi B. bassiana di lapang cenderung aman bagi musuh alami atau serangga berguna lainnya. Infeksi B. bassiana pada manusia sangat jarang terjadi. Meskipun demikian, dilaporkan ada dua kasus infeksi B. bassiana yang menyebabkan mikosis pada manusia (Henke et al., 2002; Tucker et al., 2004). Namun infeksi tersebut terjadi pada kondisi kesehatan manusia yang sangat buruk akibat penyakit leukimia akut.
Pengujian Botanigard terhadap mamalia, burung, dan ikan juga tidak menunjukkan pengaruh negatif terhadap perkembangan hewan-hewan tersebut (US EPA, 2006). Demikian pula pengujian terhadap sejumlah reptil maupun vertebrata yang membuktikan bahwa B. bassiana tidak menginfeksi keduanya (Georg et al., 1962; Fromtling et al., 1979; Gonzalez et al., 1995).
Cukup banyak tersedia bahan untuk media alami perbanyakan B. bassiana, antara lain: beras, gandum, kedelai, jagung, padi-padian, sorghum, kentang, roti, dan kacang-kacangan. Bahan mana yang akan digunakan tergantung pada beberapa faktor, termasuk kemudahan memperoleh bahan tersebut, biaya, dan strain isolat yang akan diperbanyak. Dalam perbanyakan B. bassiana dengan bahan-bahan alami, untuk menghasilkan konidia dalam jumlah maksimal diperlukan media dengan partikel yang permukaannya lebih luas. Bahan media yang cenderung menggumpal akan memiliki luas permukaan yang sempit, sehingga produksi konidia juga sedikit. Media yang ideal adalah media yang tidak hanya mempunyai partikel dengan permukaan luas, tetapi juga yang dapat mempertahankan keutuhan partikel selama proses produksi (Maheva et al., 1984; Bradley et al., 1992).
Tiga jenis bahan media alami yang telah dicoba dalam perbanyakan B. bassiana skala besar di New Zealand adalah beras, gandum, dan barley. Hasilnya, beras merupakan media paling sesuai bagi perkembangan B. bassiana dengan produktivitas konidia tertinggi mencapai 4,38 x 109 konidia/g beras (Nelson dan Glare, 1996). Penggunaan berbagai jenis sereal, selain beras, sebagai media perbanyakan B. bassiana perlu dipertimbangkan mengingat kandungan nutrisinya yang sangat bervariasi (Jenkins et al., 1998). Perbedaan kandungan nutrisi ini sangat mempengaruhi produksi konidia, terutama per kelompok produksi (batch). Oleh karena itu, pemilihan bahan media perbanyakan harus dilakukan secara tepat, terutama memilih bahan yang memiliki kemampuan produksi konidia secara konsisten dalam kelompok-kelompok produksi. Hasil penelitian lain juga membuktikan bahwa beras putih merupakan bahan media perbanyakan B. bassiana yang tepat karena produksi konidia yang tinggi (Alves dan Pereira, 1989; Mendonca, 1992; Ibrahim dan Low, 1993; Milner et al., 1993). Hal tersebut
Menunjukkan bahwa kombinasi faktor-faktor produkai sangat kompatibel, termasuk keseimbangan nutrisi dalam bahan media, biaya produksi, kemudahan memperoleh bahan, karakter fisik bahan, seperti ukuran, bentuk, dan keutuhan bahan baik sebelum maupun setelah pengkolonisasian konidia.
Dalam perbanyakan, temperatur inkubasi dan cahaya sangat menentukan produktivitas konidia. Temperatur optimal setiap cendawan bervariasi tidak saja antar spesies, tetapi juga antar isolat (Thomas dan Jenkins, 1997; Alasoadura, 1963). Temperatur optimal untuk perkecambahan konidia adalah 25-30°C, dengan temperatur minimum 10°C dan maksimum 32°C. Sedangkan pH optimal untuk pertumbuhan adalah 5,7-5,9, tapi idealnya pH 7-8 (Goral dan Lappa, 1972). Beberapa cendawan membutuhkan cahaya untuk proses sporulasi, sedangkan cendawan lainnya tidak terpengaruh oleh cahaya. Tetapi ada pula cendawan yang sporulasinya terhambat pada tingkat intensitas cahaya tertentu (Vouk dan Klas, 1931). Penelitian terdahulu membuktikan bahwa B. bassiana yang diproduksi di lingkungan tanpa cahaya (gelap) konidianya cenderung berukuran lebih besar dan lebih virulen dibanding yang diproduksi pada tempat terang (Humphreys et al., 1989; Williams, 1959). Hal ini penting sebagai bahan pertimbangan dalam memilih kemasan yang sesuai apabila biakan cendawan harus dibawa ke luar areal perbanyakan. Selain itu yang lebih penting dalam perbanyakan B. bassiana untuk skala komersial adalah kesesuaian produk dengan teknik formulasi dan aplikasinya.
Umumnya produk B. bassiana diformulasi dalam bentuk bubuk (powder) dan merupakan formulasi paling efektif memicu kontak dengan hama sasaran (Stimac et al., 1993). Formulasi B. bassiana berupa pellet hasil enkapsulasi miselium selain efektif untuk meningkatkan mortalitas hama juga untuk mengurangi kompetisi dengan mikroba lain, sehingga meningkatkan daya hidup B. bassiana (White, 1995).
Dalam jurnal diketahui kalau penambahan tepung tapioka sebanyak 1 g dan suhu penyimpanan 5OC, mempunyai potensi yang baik dalam mempertahankan viabilitas spora B. bassiana sekurang-kurangnya sampai dua bulan penyimpanan. Hal tersebut disebabkan oleh kemampuan daya simpan tepung tapioka yang lebih baik daripada tepung beras dan maizena serta didukung oleh suhu rendah yang sesuai untuk mempertahankan viabilitas spora B. bassiana. Terjadi penurunan viabilitas spora kering B. bassiana lebih cepat, seiring dengan semakin meningkatnya dosis pembawa, suhu dan lama penyimpanan. Penyimpanan pada suhu 5OC menunjukkan kemampuan mempertahankan viabilitas spora B. bassiana kering murni lebih lama daripada kondisi suhu 23C dan suhu 29C.
II. PENUTUP
Kesimpulan
Dari pembahasan di atas kita dapat menyimpulkan bahwa:
Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat
Ada beberapa faktor yang mempengaruhi mekanisme infeksi cendawan entomopatogen Beauveria bassiana, faktor yang meliputi diantaranya kelembaban, suhu, iklim, dan tingkat kekebalan tubuh dari inang
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna.
DAFTAR PUSTAKA
Anonim, 2012. Perbanyakan Cendawan Entomopatogen. http: www. Pengendalian%agents-hayati.com.html.diakses pada tanggal 21 maret 2012
Anonim, 2012. Prospek Beauveria bassiana sebagai bioinsectisida. http//:www. Miss Biology Prospek Beauveria bassiana sebagai bioinsectisida.htm. diakses pada tanggal 21 maret 2012
Anonim, 2012. Beauveria bassiana . Wikipedia bahasa Indonesia, ensiklopedia bebas.html. diakses pada tanggal 21 maret 2012
Balai Penelitian Tanaman Hias. 2006. Beauveria bassiana pengendali hama tanaman. Warta Penelitian dan Pengembangan Pertanian Indonesia 28(1): 11-12.
EOL. 2000. Beauveria bassiana Bals. -Criv. Vuill. 1912.[terhubung berkala] http://www.eol.org/pages/160292 . diakses pada tanggal 21 maret 2012
McCoy C, Quintela ED, Faria M de. 1990. Environmental persistence of entomopathogenic fungi. [terhubung berkala]. http://www.lsuagcenter.com/s265/mccoy.htm. diakses pada tanggal 21 maret 2012
Agens Pengendali Hayati adalah musuh alami berupa Parasitoid, Predator, Entomopatogen dan Agens Antagonis yang telah diketahui mempunyai potensi yang besar sebagai salah satu sarana pengendalian OPT.
Entomopatogen atau penyakit pada serangga hama yang mencakup semua mikroba yaitu Cendawan, bakteri, virus, dan nematoda yang memiliki kemampuan untuk menjadi parasit (pathogen) pada serangga sehingga dapat mematikan serangga yang diinfeksinya. Cendawan adalah mikroba yang mempunyai bentuk tubuh menyerupai benang yang disebut hifa, nampak seperti tumbuhan namun tidak memiliki klorofil.
Mikroba dari jenis jamur adalah Beauveria bassiana, Metarrhizium anisopliae Metarrhizium ftavoviridae Verticillium lecanii dll. dapat mengendalikan Uret, Kepinding tanah, Belalang, Lalat buah dan serangga lainnya.
Kesulitan dan permasalahan utama dalam penerapan dan pengembangan pengendalian hayati adalah modal investasi permulaan yang besar yang harus dikeluarkan untuk kegiatan eksplorasi, penelitian, pengujian dan evaluasi terutama yang menyangkut berbagai aspek dasar baik untuk hama, musuh alami maupun tanaman. Aspek dasar dapat meliputi taksonomi, ekologi, biologi, siklus hidup, dinamika populasi, genetika, fisiologi, dll. Identifikasi yang tepat baik untuk jenis hama maupun musuh alaminya merupakan langkah permulaan yang sangat penting.
Kecuali diperlukan modal, fasilitas yang lengkap juga diperlukan sumber daya manusia terutama para peneliti yang berkualitas dan berpendidikan khusus dan berdedikasi tinggi sesuai dengan yang diperlukan untuk pengembangan teknologi pengendalian hayati. Meskipun ada beberapa ahli yang berpendapat bahwa untuk pengendalian hayati yang penting adalah adanya tenaga peneliti yang berpengalaman dan berdedikasi tinggi serta cukup memiliki rasa seni dan intuisi, namun bagaimanapun untuk keberhasilan pengendalian hayati dalam kerangka PHT diperlukan juga dasar pengetahuan dan teknologi yang mantap.
II. PEMBAHASAN
Adanya kekhawatiran akan pengaruh negatif tentang pemakaian agrokhemikal telah meningkatkan perhatian masyarakat kepada bioinsektisida sebagai alternative teknologi untuk menurunkan populasi hama. Salah satu alternatif pengendalian adalah pemanfaatan jamur penyebab penyakit pada serangga (bioinsectisida), yaitu jamur patogen serangga Beauveria bassiana.
Di Indonesia, hasil-hasil penelitian B. bassiana juga telah banyak dipublikasikan, terutama dari tanaman pangan untuk mengendalikan serangga hama kedelai (Riptortus linearis dan Spodoptera litura), walang sangit pada padi (Leptocoriza acuta) (Prayogo, 2006), Plutella xylostella pada sayursayuran (Hardiyanti, 2006), hama bubuk buah kopi Helopeltis antoni, dan penggerek buah kakaoHypothenemus hampei (Sudarmadji dan Prayogo, dalam Prayogo, 2006).
Saat ini produk bioinsektisida berbahan aktif B. bassiana telah tersedia secara komersial di Indonesia. Meskipun demikian, tampaknya pemanfaatannya di lapang khususnya untuk tanaman perkebunan belum optimal. Padahal,lingkungan mikro tanaman perkebunan sangat ideal bagi perkembangan epizootik cendawan-cendawan entomopatogen, termasuk B. bassiana. Keberlangsungan epizootik cendawan sangat dipengaruhi oleh suhu dan kelembaban lingkungan, dan kriteria ini dapat ditemukan pada tanaman-tanaman perkebunan yang banyak diusahakan di Indonesia. Disamping itu, pemanfaatan cendawan ini dan patogen serangga secara umum dalam pengendalian hama berpotensi memberi keuntungan ekologis jangka panjang terhadap keseimbangan hayati maupun keberlanjutan sistem pertanian.
Pengendalian penyakit tanaman dengan menggunakan agens pengendali hayati muncul karena kekhawatiran masyarakat dunia akibat penggunaan pestisida kimia sintetis. Adanya kekhawatiran tersebut membuat pengendalian hayati menjadi salah satu pilihan cara mengendalikan patogen tanaman yang harus dipertimbangkan.
Agensia hayati adalah setiap organisme yang meliputi spesies, subspesies, varietas, semua jenis serangga, nematoda, protozoa, cendawan (fungi), bakteri, virus, mikoplasma, serta organisme lainnya dalam semua tahap perkembangannya yang dapat dipergunakan untuk keperluan pengendalian hama dan penyakit atau organisme pengganggu, proses produksi, pengolahan hasil pertanian, dan berbagai keperluan lainnya (Permentan No. 411 tahun 1995).
Beauvaria bassiana merupakan cendawan entomopatogen yaitu cendawan yang dapat menimbulkan penyakit pada serangga. Beauveria bassiana secara alami terdapat di dalam tanah sebagai jamur saprofit. Pertumbuhan jamur di dalam tanah sangat dipengaruhi oleh kondisi tanah, seperti kandungan bahan organik, suhu, kelembapan, kebiasaan makan serangga, adanya pestisida sintetis, dan waktu aplikasi. Secara umum, suhu di atas 30 °C, kelembapan tanah yang berkurang dan adanya antifungal atau pestisida dapat menghambat pertumbuhannya.
Beauveria bassiana membunuh hama melalui infeksi sebagai akibat dari serangga yang kontak dengan spora jamur. Serangga dapat kontak dengan spora jamur melalui beberapa cara: semprotan jamur menempel pada tubuh serangga, serangga bergerak pada permukaan tanaman yang sudah terinfeksi jamur, atau dengan memakan jaringan tanaman yang telah diperlakukan dengan jamur. Setelah spora jamur melekat pada kulit serangga (kutikula), mereka berkecambah membentuk struktur (hifa) yang menembus tubuh serangga dan berkembang biak. Proses ini memakan waktu 3-5 hari sampai akhirnya serangga mati, bangkai yang terinfeksi dapat berfungsi sebagai sumber spora untuk penyebaran sekunder jamur. Keberhasilan penyemprotan menggunakan jamur Beauveria bassiana bergantung pada kerentanan spesies yang bersangkutan, tingkat populasi hama, dan kondisi lingkungan pada saat aplikasi, serta sumber daya manusia itu sendiri.
Mekanisme Infeksi Beauveria Bassiana
B. bassiana memproduksi toksin yang disebut beauvericin (Kučera dan Samšiňáková, 1968). Antibiotik ini dapat menyebabkan gangguan pada fungsi hemolimfa dan nukleus serangga, sehingga mengakibatkan pembengkakan yang disertai pengerasan pada serangga yang terinfeksi. Selain secara kontak, B. bassiana juga dapat menginfeksi serangga melalui inokulasi atau kontaminasi pakan. Broome et al. (1976) menyatakan bahwa 37% dari konidia B. bassiana yang dicampurkan ke dalam pakan semut api, Selenopsis richteri, berkecambah di dalam saluran pencernaan inangnya dalam waktu 72 jam, sedangkan hifanya mampu menembus dinding usus antara 60-72 jam. Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat. Serangga yang mati tidak selalu disertai gejala pertumbuhan spora. Contohnya, aphid yang terinfeksi B.bassiana hanya mengalami pembengkakan tanpa terjadi perubahan warna. Demikian pula tempayak lalat yang terinfeksi B. bassiana sering ditemukan secara berkelompok pada ujung-ujung rerumputan (Plate, 1976).
Kematian serangga biasanya disebabkan oleh kerusakan jaringan secara menyeluruh, dan atau karena toksin yang diproduksi oleh cendawan. Menurut Cheung dan Grula (1982), penyakit white muscardine yang menyerang saluran pencernaan Heliothis zea mengakibatkan gangguan nutrisi hingga kematian. Serangga yang terbunuh tubuhnya akan berwarna putih karena ditumbuhi konidia B. bassiana. Jumlah konidia yang dapat dihasilkan oleh satu serangga ditentukan oleh besar kecilnya ukuran serangga tersebut. Setiap serangga terinfeksi B. bassiana akan efektif menjadi sumber infeksi bagi serangga sehat di sekitarnya.
Seperti cendawan lain, perrtumbuhan B. bassiana juga sangat ditentukan oleh kelembapan lingkungan. Namun demikian, cendawan ini juga memiliki fase resisten yang dapat mempertahankan kemampuannya menginfeksi inang pada kondisi kering. Keberadaan epizootiknya di alam menyebabkan B. bassiana secara cepat menginfeksi populasi serangga hingga menyebabkan kematian. Selain itu, kemampuan penetrasinya yang tinggi pada tubuh serangga menyebabkan cendawan ini juga dengan mudah menginfeksi serangga hama pengisap, seperti aphid (Aphis sp.) dan kutu putih Bemisia spp. yang tidak mudah terinfeksi oleh bakteri maupun virus.
Faktor lingkungan, terutama kelembaban dan temperatur serta sedikit cahaya sangat penting perannya dalam proses infeksi dan sporulasi cendawan entomopatogen (Roberts dan Campbell, 1977; McCoy et al., 1988). Temperatur optimum untuk perkembangan, patogenisitas, dan kelulusan hidup cendawan umumnya antara 20-30°C (McCoy et al., 1988). Untuk perkecambahan konidia dan sporulasi pada permukaan tubuh serangga dibutuhkan kelembaban sangat tinggi (> 90% RH), terutama kelembaban di lingkungan mikro sekitar konidia sangat penting perannya dalam proses perkecambahan dan produksi konidia (Millstein et al., 1983; Nordin et al., 1983). Tetapi sebaliknya untuk melepaskan konidia B. bassiana dari konidifor hanya dibutuhkan kelembaban sekitar 50% (Gottwald dan Tedders, 1982).
Meskipun pengaruh cahaya terhadap infeksi cendawan belum diketahui secara jelas, tetapi intensitas sinar ultraviolet tertentu dapat merusak konidia cendawan (Callaghan, 1969). Fuxa (1987) menyatakan bahwa intensitas cahaya matahari dengan rata-rata panjang gelombang antara 290-400 nm cukup efektif menurunkan persistensi deposit konidia pada pertanaman. Sementara Ignoffo et al. (1977) mengemukakan bahwa waktu paruh (half-life) sebagian besar spora cendawan yang terekspos cahaya buatan dengan panjang gelombang mendekati panjang gelombang sinar matahari (290-400 nm) hanya sekitar 1-4 jam, tetapi kenyataannya di lapang waktu paruh dapat mencapai lebih dari 4 jam.
Umumnya cendawan entomopatogen membutuhkan lingkungan yang lembab untuk dapat menginfeksi serangga, oleh karena itu epizootiknya di alam biasanya terbentuk pada saat kondisi lingkungan lembab atau basah. Keefektifan B. bassiana menginfeksi serangga hama tergantung pada spesies atau strain cendawan, dan kepekaan stadia serangga pada tingkat kelembaban lingkungan, struktur tanah (untuk serangga dalam tanah), dan temperatur yang tepat. Selain itu, harus terjadi kontak antara spora B. bassiana yang diterbangkan angin atau terbawa air dengan serangga inang agar terjadi infeksi. Plate (1976) juga menyatakan bahwa epizootik cendawan yang terbentuk secara alami efektif mengendalikan populasi aphid, tempayak lalat yang menyerang perakaran tanaman, belalang, dan thrip, disamping juga potensial sebagai faktor mortalitas utama aphid yang menyerang kentang dan tanaman inang lainnya.
Konidia merupakan unit B. bassiana yang paling infektif dan stabil untuk aplikasi di lapang dibandingkan dengan hifa maupun blastosporanya (Soper dan Ward, 1981; Feng et al., 1994). Konidia yang diaplikasikan dapat berupa suspensi (tidak diformulasi), formulasi butiran, dan bentuk pellet, dan ketiganya memperlihatkan hasil pengendalian yang cukup nyata. Stimac et al. (1993) menyatakan bahwa aplikasi konidia B. bassiana dengan cara sprinkle dan disemprotkan pada permukaan tanah sangat efektif menyebabkan mortalitas hama sasaran. Mortalitas hama semut api, Selenopsis invicta, yang dikendalikan dengan B. bassiana tertinggi mulai 3-8 hari setelah perlakuan. Sedangkan enkapsulasi (pellet) konidia B. bassiana dengan menggunakan kalsium alginat juga efektif meningkatkan mortalitas S. invicta (White, 1995), karena enkapsulasi menyebabkan konidia lebih stabil di dalam tanah.
Beberapa senyawa metabolit sekunder diproduksi oleh B. bassiana, seperti beauvericin, bassianin, bassiacridin, bassianolide, beauverolides, tenellin, dan oosporein (Strasser et al., 2000; Vey et al., 2001; Quesada-Moraga dan Vey, 2004). Senyawa metabolit sekunder ini dapat dihasilkan oleh B. bassiana pada epizootik di alam (tanah) maupun pada epizootik buatan (di laboratorium). Meskipun demikian, hingga saat ini belum ada laporan tentang tercemarnya rantai makanan oleh senyawa metabolit sekunder, atau terakumulasi di alam sebagai limbah epizootik B. bassiana (Vey et al., 2001).
Penggunaan B. bassiana dalam pengendalian hama telah diuji secara luas di berbagai negara. Hasil uji toksikologi terhadap salah satu produk B. bassiana, Botanigard, menunjukkan bahwa produk tersebut tidak menimbulkan dampak negatif yang berhubungan dengan patogenisitas dan toksisitasnya, sehingga produk tersebut digunakan secara aman selama lebih dari 10 tahun di Amerika Serikat dan juga di beberapa negara lain (US EPA, 2006).
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna. Namun, Plate (1976) mengungkapkan bahwa tingkat kepekaan serangga bukan sasaran terhadap infeksi B. bassiana sangat ditentukan oleh virulensi dan patogenisitas cendawan, serta spesies serangga inang. Selain itu, perbedaan fisiologis dan ekologis inang juga mempengaruhi infeksi B. bassiana. Misalnya, serangga bukan sasaran yang mudah terinfeksi B. bassiana di laboratorium tidak akan serta merta terinfeksi pada kondisi lapang. Ludwig dan Oetting (2001) menegaskan bahwa beberapa serangga musuh alami yang peka terhadap infeksi B. bassiana di laboratorium ternyata mengalami infeksi sangat rendah pada uji di rumah kaca. Disamping itu, hasil uji ekotoksikologi terhadap produk Botanigard menunjukkan bahwa risiko secara ekologis yang diperlihatkan oleh serangga bukan sasaran yang diperlakukan dengan formulasi B. bassiana sangat rendah (US EPA, 2006).
Dihubungkan dengan keamanan secara hayati, cendawan entomopatogen dikelompokkan menjadi cendawan dengan kisaran inang spesifik dan yang kisaran inangnya luas (MacLeod, 1963). Cendawan yang memiliki kisaran inang spesifik umumnya menjadi parasit sejati (obligat) dan bersifat sangat virulen tarhadap inang. Sebaliknya yang kisaran inangnya luas sebagian besar merupakan patogen fakultatif, bersifat saprofit, dan cenderung kurang patogenik (Goettel et al., 1990), dan biasanya virulensinya tinggi hanya pada spesies inang dari mana cendawan tersebut pertama kali diisolasi. Contoh, B. bassiana yang diisolasi dari ulat H. armigera akan lebih patogenik pada inangnya tersebut dibanding dengan inang-inangnya yang lain. Selain itu, cendawan yang kisaran inangnya lebih luas justru menjadi lebih spesifik menginfeksi inang jika di lapang. Hal ini dapat terjadi kemungkinan karena dipengaruhi oleh interaksi antara faktor abiotik dan biotik di lapang, sehingga serangga yang mudah terinfeksi di laboratorium belum tentu mudah juga terinfeksi di lapang. Oleh karena itu, kemungkinan terinfeksinya serangga bukan sasaran oleh B. bassiana di lapang sangat kecil. Dengan demikian, aplikasi B. bassiana di lapang cenderung aman bagi musuh alami atau serangga berguna lainnya. Infeksi B. bassiana pada manusia sangat jarang terjadi. Meskipun demikian, dilaporkan ada dua kasus infeksi B. bassiana yang menyebabkan mikosis pada manusia (Henke et al., 2002; Tucker et al., 2004). Namun infeksi tersebut terjadi pada kondisi kesehatan manusia yang sangat buruk akibat penyakit leukimia akut.
Pengujian Botanigard terhadap mamalia, burung, dan ikan juga tidak menunjukkan pengaruh negatif terhadap perkembangan hewan-hewan tersebut (US EPA, 2006). Demikian pula pengujian terhadap sejumlah reptil maupun vertebrata yang membuktikan bahwa B. bassiana tidak menginfeksi keduanya (Georg et al., 1962; Fromtling et al., 1979; Gonzalez et al., 1995).
Cukup banyak tersedia bahan untuk media alami perbanyakan B. bassiana, antara lain: beras, gandum, kedelai, jagung, padi-padian, sorghum, kentang, roti, dan kacang-kacangan. Bahan mana yang akan digunakan tergantung pada beberapa faktor, termasuk kemudahan memperoleh bahan tersebut, biaya, dan strain isolat yang akan diperbanyak. Dalam perbanyakan B. bassiana dengan bahan-bahan alami, untuk menghasilkan konidia dalam jumlah maksimal diperlukan media dengan partikel yang permukaannya lebih luas. Bahan media yang cenderung menggumpal akan memiliki luas permukaan yang sempit, sehingga produksi konidia juga sedikit. Media yang ideal adalah media yang tidak hanya mempunyai partikel dengan permukaan luas, tetapi juga yang dapat mempertahankan keutuhan partikel selama proses produksi (Maheva et al., 1984; Bradley et al., 1992).
Tiga jenis bahan media alami yang telah dicoba dalam perbanyakan B. bassiana skala besar di New Zealand adalah beras, gandum, dan barley. Hasilnya, beras merupakan media paling sesuai bagi perkembangan B. bassiana dengan produktivitas konidia tertinggi mencapai 4,38 x 109 konidia/g beras (Nelson dan Glare, 1996). Penggunaan berbagai jenis sereal, selain beras, sebagai media perbanyakan B. bassiana perlu dipertimbangkan mengingat kandungan nutrisinya yang sangat bervariasi (Jenkins et al., 1998). Perbedaan kandungan nutrisi ini sangat mempengaruhi produksi konidia, terutama per kelompok produksi (batch). Oleh karena itu, pemilihan bahan media perbanyakan harus dilakukan secara tepat, terutama memilih bahan yang memiliki kemampuan produksi konidia secara konsisten dalam kelompok-kelompok produksi. Hasil penelitian lain juga membuktikan bahwa beras putih merupakan bahan media perbanyakan B. bassiana yang tepat karena produksi konidia yang tinggi (Alves dan Pereira, 1989; Mendonca, 1992; Ibrahim dan Low, 1993; Milner et al., 1993). Hal tersebut
Menunjukkan bahwa kombinasi faktor-faktor produkai sangat kompatibel, termasuk keseimbangan nutrisi dalam bahan media, biaya produksi, kemudahan memperoleh bahan, karakter fisik bahan, seperti ukuran, bentuk, dan keutuhan bahan baik sebelum maupun setelah pengkolonisasian konidia.
Dalam perbanyakan, temperatur inkubasi dan cahaya sangat menentukan produktivitas konidia. Temperatur optimal setiap cendawan bervariasi tidak saja antar spesies, tetapi juga antar isolat (Thomas dan Jenkins, 1997; Alasoadura, 1963). Temperatur optimal untuk perkecambahan konidia adalah 25-30°C, dengan temperatur minimum 10°C dan maksimum 32°C. Sedangkan pH optimal untuk pertumbuhan adalah 5,7-5,9, tapi idealnya pH 7-8 (Goral dan Lappa, 1972). Beberapa cendawan membutuhkan cahaya untuk proses sporulasi, sedangkan cendawan lainnya tidak terpengaruh oleh cahaya. Tetapi ada pula cendawan yang sporulasinya terhambat pada tingkat intensitas cahaya tertentu (Vouk dan Klas, 1931). Penelitian terdahulu membuktikan bahwa B. bassiana yang diproduksi di lingkungan tanpa cahaya (gelap) konidianya cenderung berukuran lebih besar dan lebih virulen dibanding yang diproduksi pada tempat terang (Humphreys et al., 1989; Williams, 1959). Hal ini penting sebagai bahan pertimbangan dalam memilih kemasan yang sesuai apabila biakan cendawan harus dibawa ke luar areal perbanyakan. Selain itu yang lebih penting dalam perbanyakan B. bassiana untuk skala komersial adalah kesesuaian produk dengan teknik formulasi dan aplikasinya.
Umumnya produk B. bassiana diformulasi dalam bentuk bubuk (powder) dan merupakan formulasi paling efektif memicu kontak dengan hama sasaran (Stimac et al., 1993). Formulasi B. bassiana berupa pellet hasil enkapsulasi miselium selain efektif untuk meningkatkan mortalitas hama juga untuk mengurangi kompetisi dengan mikroba lain, sehingga meningkatkan daya hidup B. bassiana (White, 1995).
Dalam jurnal diketahui kalau penambahan tepung tapioka sebanyak 1 g dan suhu penyimpanan 5OC, mempunyai potensi yang baik dalam mempertahankan viabilitas spora B. bassiana sekurang-kurangnya sampai dua bulan penyimpanan. Hal tersebut disebabkan oleh kemampuan daya simpan tepung tapioka yang lebih baik daripada tepung beras dan maizena serta didukung oleh suhu rendah yang sesuai untuk mempertahankan viabilitas spora B. bassiana. Terjadi penurunan viabilitas spora kering B. bassiana lebih cepat, seiring dengan semakin meningkatnya dosis pembawa, suhu dan lama penyimpanan. Penyimpanan pada suhu 5OC menunjukkan kemampuan mempertahankan viabilitas spora B. bassiana kering murni lebih lama daripada kondisi suhu 23C dan suhu 29C.
II. PENUTUP
Kesimpulan
Dari pembahasan di atas kita dapat menyimpulkan bahwa:
Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat
Ada beberapa faktor yang mempengaruhi mekanisme infeksi cendawan entomopatogen Beauveria bassiana, faktor yang meliputi diantaranya kelembaban, suhu, iklim, dan tingkat kekebalan tubuh dari inang
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna.
DAFTAR PUSTAKA
Anonim, 2012. Perbanyakan Cendawan Entomopatogen. http: www. Pengendalian%agents-hayati.com.html.diakses pada tanggal 21 maret 2012
Anonim, 2012. Prospek Beauveria bassiana sebagai bioinsectisida. http//:www. Miss Biology Prospek Beauveria bassiana sebagai bioinsectisida.htm. diakses pada tanggal 21 maret 2012
Anonim, 2012. Beauveria bassiana . Wikipedia bahasa Indonesia, ensiklopedia bebas.html. diakses pada tanggal 21 maret 2012
Balai Penelitian Tanaman Hias. 2006. Beauveria bassiana pengendali hama tanaman. Warta Penelitian dan Pengembangan Pertanian Indonesia 28(1): 11-12.
EOL. 2000. Beauveria bassiana Bals. -Criv. Vuill. 1912.[terhubung berkala] http://www.eol.org/pages/160292 . diakses pada tanggal 21 maret 2012
McCoy C, Quintela ED, Faria M de. 1990. Environmental persistence of entomopathogenic fungi. [terhubung berkala]. http://www.lsuagcenter.com/s265/mccoy.htm. diakses pada tanggal 21 maret 2012
I. PENDAHULUAN
Agens Pengendali Hayati adalah musuh alami berupa Parasitoid, Predator, Entomopatogen dan Agens Antagonis yang telah diketahui mempunyai potensi yang besar sebagai salah satu sarana pengendalian OPT.
Entomopatogen atau penyakit pada serangga hama yang mencakup semua mikroba yaitu Cendawan, bakteri, virus, dan nematoda yang memiliki kemampuan untuk menjadi parasit (pathogen) pada serangga sehingga dapat mematikan serangga yang diinfeksinya. Cendawan adalah mikroba yang mempunyai bentuk tubuh menyerupai benang yang disebut hifa, nampak seperti tumbuhan namun tidak memiliki klorofil.
Mikroba dari jenis jamur adalah Beauveria bassiana, Metarrhizium anisopliae Metarrhizium ftavoviridae Verticillium lecanii dll. dapat mengendalikan Uret, Kepinding tanah, Belalang, Lalat buah dan serangga lainnya.
Kesulitan dan permasalahan utama dalam penerapan dan pengembangan pengendalian hayati adalah modal investasi permulaan yang besar yang harus dikeluarkan untuk kegiatan eksplorasi, penelitian, pengujian dan evaluasi terutama yang menyangkut berbagai aspek dasar baik untuk hama, musuh alami maupun tanaman. Aspek dasar dapat meliputi taksonomi, ekologi, biologi, siklus hidup, dinamika populasi, genetika, fisiologi, dll. Identifikasi yang tepat baik untuk jenis hama maupun musuh alaminya merupakan langkah permulaan yang sangat penting.
Kecuali diperlukan modal, fasilitas yang lengkap juga diperlukan sumber daya manusia terutama para peneliti yang berkualitas dan berpendidikan khusus dan berdedikasi tinggi sesuai dengan yang diperlukan untuk pengembangan teknologi pengendalian hayati. Meskipun ada beberapa ahli yang berpendapat bahwa untuk pengendalian hayati yang penting adalah adanya tenaga peneliti yang berpengalaman dan berdedikasi tinggi serta cukup memiliki rasa seni dan intuisi, namun bagaimanapun untuk keberhasilan pengendalian hayati dalam kerangka PHT diperlukan juga dasar pengetahuan dan teknologi yang mantap.
II. PEMBAHASAN
Adanya kekhawatiran akan pengaruh negatif tentang pemakaian agrokhemikal telah meningkatkan perhatian masyarakat kepada bioinsektisida sebagai alternative teknologi untuk menurunkan populasi hama. Salah satu alternatif pengendalian adalah pemanfaatan jamur penyebab penyakit pada serangga (bioinsectisida), yaitu jamur patogen serangga Beauveria bassiana.
Di Indonesia, hasil-hasil penelitian B. bassiana juga telah banyak dipublikasikan, terutama dari tanaman pangan untuk mengendalikan serangga hama kedelai (Riptortus linearis dan Spodoptera litura), walang sangit pada padi (Leptocoriza acuta) (Prayogo, 2006), Plutella xylostella pada sayursayuran (Hardiyanti, 2006), hama bubuk buah kopi Helopeltis antoni, dan penggerek buah kakaoHypothenemus hampei (Sudarmadji dan Prayogo, dalam Prayogo, 2006).
Saat ini produk bioinsektisida berbahan aktif B. bassiana telah tersedia secara komersial di Indonesia. Meskipun demikian, tampaknya pemanfaatannya di lapang khususnya untuk tanaman perkebunan belum optimal. Padahal,lingkungan mikro tanaman perkebunan sangat ideal bagi perkembangan epizootik cendawan-cendawan entomopatogen, termasuk B. bassiana. Keberlangsungan epizootik cendawan sangat dipengaruhi oleh suhu dan kelembaban lingkungan, dan kriteria ini dapat ditemukan pada tanaman-tanaman perkebunan yang banyak diusahakan di Indonesia. Disamping itu, pemanfaatan cendawan ini dan patogen serangga secara umum dalam pengendalian hama berpotensi memberi keuntungan ekologis jangka panjang terhadap keseimbangan hayati maupun keberlanjutan sistem pertanian.
Pengendalian penyakit tanaman dengan menggunakan agens pengendali hayati muncul karena kekhawatiran masyarakat dunia akibat penggunaan pestisida kimia sintetis. Adanya kekhawatiran tersebut membuat pengendalian hayati menjadi salah satu pilihan cara mengendalikan patogen tanaman yang harus dipertimbangkan.
Agensia hayati adalah setiap organisme yang meliputi spesies, subspesies, varietas, semua jenis serangga, nematoda, protozoa, cendawan (fungi), bakteri, virus, mikoplasma, serta organisme lainnya dalam semua tahap perkembangannya yang dapat dipergunakan untuk keperluan pengendalian hama dan penyakit atau organisme pengganggu, proses produksi, pengolahan hasil pertanian, dan berbagai keperluan lainnya (Permentan No. 411 tahun 1995).
Beauvaria bassiana merupakan cendawan entomopatogen yaitu cendawan yang dapat menimbulkan penyakit pada serangga. Beauveria bassiana secara alami terdapat di dalam tanah sebagai jamur saprofit. Pertumbuhan jamur di dalam tanah sangat dipengaruhi oleh kondisi tanah, seperti kandungan bahan organik, suhu, kelembapan, kebiasaan makan serangga, adanya pestisida sintetis, dan waktu aplikasi. Secara umum, suhu di atas 30 °C, kelembapan tanah yang berkurang dan adanya antifungal atau pestisida dapat menghambat pertumbuhannya.
Beauveria bassiana membunuh hama melalui infeksi sebagai akibat dari serangga yang kontak dengan spora jamur. Serangga dapat kontak dengan spora jamur melalui beberapa cara: semprotan jamur menempel pada tubuh serangga, serangga bergerak pada permukaan tanaman yang sudah terinfeksi jamur, atau dengan memakan jaringan tanaman yang telah diperlakukan dengan jamur. Setelah spora jamur melekat pada kulit serangga (kutikula), mereka berkecambah membentuk struktur (hifa) yang menembus tubuh serangga dan berkembang biak. Proses ini memakan waktu 3-5 hari sampai akhirnya serangga mati, bangkai yang terinfeksi dapat berfungsi sebagai sumber spora untuk penyebaran sekunder jamur. Keberhasilan penyemprotan menggunakan jamur Beauveria bassiana bergantung pada kerentanan spesies yang bersangkutan, tingkat populasi hama, dan kondisi lingkungan pada saat aplikasi, serta sumber daya manusia itu sendiri.
Mekanisme Infeksi Beauveria Bassiana
B. bassiana memproduksi toksin yang disebut beauvericin (Kučera dan Samšiňáková, 1968). Antibiotik ini dapat menyebabkan gangguan pada fungsi hemolimfa dan nukleus serangga, sehingga mengakibatkan pembengkakan yang disertai pengerasan pada serangga yang terinfeksi. Selain secara kontak, B. bassiana juga dapat menginfeksi serangga melalui inokulasi atau kontaminasi pakan. Broome et al. (1976) menyatakan bahwa 37% dari konidia B. bassiana yang dicampurkan ke dalam pakan semut api, Selenopsis richteri, berkecambah di dalam saluran pencernaan inangnya dalam waktu 72 jam, sedangkan hifanya mampu menembus dinding usus antara 60-72 jam. Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat. Serangga yang mati tidak selalu disertai gejala pertumbuhan spora. Contohnya, aphid yang terinfeksi B.bassiana hanya mengalami pembengkakan tanpa terjadi perubahan warna. Demikian pula tempayak lalat yang terinfeksi B. bassiana sering ditemukan secara berkelompok pada ujung-ujung rerumputan (Plate, 1976).
Kematian serangga biasanya disebabkan oleh kerusakan jaringan secara menyeluruh, dan atau karena toksin yang diproduksi oleh cendawan. Menurut Cheung dan Grula (1982), penyakit white muscardine yang menyerang saluran pencernaan Heliothis zea mengakibatkan gangguan nutrisi hingga kematian. Serangga yang terbunuh tubuhnya akan berwarna putih karena ditumbuhi konidia B. bassiana. Jumlah konidia yang dapat dihasilkan oleh satu serangga ditentukan oleh besar kecilnya ukuran serangga tersebut. Setiap serangga terinfeksi B. bassiana akan efektif menjadi sumber infeksi bagi serangga sehat di sekitarnya.
Seperti cendawan lain, perrtumbuhan B. bassiana juga sangat ditentukan oleh kelembapan lingkungan. Namun demikian, cendawan ini juga memiliki fase resisten yang dapat mempertahankan kemampuannya menginfeksi inang pada kondisi kering. Keberadaan epizootiknya di alam menyebabkan B. bassiana secara cepat menginfeksi populasi serangga hingga menyebabkan kematian. Selain itu, kemampuan penetrasinya yang tinggi pada tubuh serangga menyebabkan cendawan ini juga dengan mudah menginfeksi serangga hama pengisap, seperti aphid (Aphis sp.) dan kutu putih Bemisia spp. yang tidak mudah terinfeksi oleh bakteri maupun virus.
Faktor lingkungan, terutama kelembaban dan temperatur serta sedikit cahaya sangat penting perannya dalam proses infeksi dan sporulasi cendawan entomopatogen (Roberts dan Campbell, 1977; McCoy et al., 1988). Temperatur optimum untuk perkembangan, patogenisitas, dan kelulusan hidup cendawan umumnya antara 20-30°C (McCoy et al., 1988). Untuk perkecambahan konidia dan sporulasi pada permukaan tubuh serangga dibutuhkan kelembaban sangat tinggi (> 90% RH), terutama kelembaban di lingkungan mikro sekitar konidia sangat penting perannya dalam proses perkecambahan dan produksi konidia (Millstein et al., 1983; Nordin et al., 1983). Tetapi sebaliknya untuk melepaskan konidia B. bassiana dari konidifor hanya dibutuhkan kelembaban sekitar 50% (Gottwald dan Tedders, 1982).
Meskipun pengaruh cahaya terhadap infeksi cendawan belum diketahui secara jelas, tetapi intensitas sinar ultraviolet tertentu dapat merusak konidia cendawan (Callaghan, 1969). Fuxa (1987) menyatakan bahwa intensitas cahaya matahari dengan rata-rata panjang gelombang antara 290-400 nm cukup efektif menurunkan persistensi deposit konidia pada pertanaman. Sementara Ignoffo et al. (1977) mengemukakan bahwa waktu paruh (half-life) sebagian besar spora cendawan yang terekspos cahaya buatan dengan panjang gelombang mendekati panjang gelombang sinar matahari (290-400 nm) hanya sekitar 1-4 jam, tetapi kenyataannya di lapang waktu paruh dapat mencapai lebih dari 4 jam.
Umumnya cendawan entomopatogen membutuhkan lingkungan yang lembab untuk dapat menginfeksi serangga, oleh karena itu epizootiknya di alam biasanya terbentuk pada saat kondisi lingkungan lembab atau basah. Keefektifan B. bassiana menginfeksi serangga hama tergantung pada spesies atau strain cendawan, dan kepekaan stadia serangga pada tingkat kelembaban lingkungan, struktur tanah (untuk serangga dalam tanah), dan temperatur yang tepat. Selain itu, harus terjadi kontak antara spora B. bassiana yang diterbangkan angin atau terbawa air dengan serangga inang agar terjadi infeksi. Plate (1976) juga menyatakan bahwa epizootik cendawan yang terbentuk secara alami efektif mengendalikan populasi aphid, tempayak lalat yang menyerang perakaran tanaman, belalang, dan thrip, disamping juga potensial sebagai faktor mortalitas utama aphid yang menyerang kentang dan tanaman inang lainnya.
Konidia merupakan unit B. bassiana yang paling infektif dan stabil untuk aplikasi di lapang dibandingkan dengan hifa maupun blastosporanya (Soper dan Ward, 1981; Feng et al., 1994). Konidia yang diaplikasikan dapat berupa suspensi (tidak diformulasi), formulasi butiran, dan bentuk pellet, dan ketiganya memperlihatkan hasil pengendalian yang cukup nyata. Stimac et al. (1993) menyatakan bahwa aplikasi konidia B. bassiana dengan cara sprinkle dan disemprotkan pada permukaan tanah sangat efektif menyebabkan mortalitas hama sasaran. Mortalitas hama semut api, Selenopsis invicta, yang dikendalikan dengan B. bassiana tertinggi mulai 3-8 hari setelah perlakuan. Sedangkan enkapsulasi (pellet) konidia B. bassiana dengan menggunakan kalsium alginat juga efektif meningkatkan mortalitas S. invicta (White, 1995), karena enkapsulasi menyebabkan konidia lebih stabil di dalam tanah.
Beberapa senyawa metabolit sekunder diproduksi oleh B. bassiana, seperti beauvericin, bassianin, bassiacridin, bassianolide, beauverolides, tenellin, dan oosporein (Strasser et al., 2000; Vey et al., 2001; Quesada-Moraga dan Vey, 2004). Senyawa metabolit sekunder ini dapat dihasilkan oleh B. bassiana pada epizootik di alam (tanah) maupun pada epizootik buatan (di laboratorium). Meskipun demikian, hingga saat ini belum ada laporan tentang tercemarnya rantai makanan oleh senyawa metabolit sekunder, atau terakumulasi di alam sebagai limbah epizootik B. bassiana (Vey et al., 2001).
Penggunaan B. bassiana dalam pengendalian hama telah diuji secara luas di berbagai negara. Hasil uji toksikologi terhadap salah satu produk B. bassiana, Botanigard, menunjukkan bahwa produk tersebut tidak menimbulkan dampak negatif yang berhubungan dengan patogenisitas dan toksisitasnya, sehingga produk tersebut digunakan secara aman selama lebih dari 10 tahun di Amerika Serikat dan juga di beberapa negara lain (US EPA, 2006).
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna. Namun, Plate (1976) mengungkapkan bahwa tingkat kepekaan serangga bukan sasaran terhadap infeksi B. bassiana sangat ditentukan oleh virulensi dan patogenisitas cendawan, serta spesies serangga inang. Selain itu, perbedaan fisiologis dan ekologis inang juga mempengaruhi infeksi B. bassiana. Misalnya, serangga bukan sasaran yang mudah terinfeksi B. bassiana di laboratorium tidak akan serta merta terinfeksi pada kondisi lapang. Ludwig dan Oetting (2001) menegaskan bahwa beberapa serangga musuh alami yang peka terhadap infeksi B. bassiana di laboratorium ternyata mengalami infeksi sangat rendah pada uji di rumah kaca. Disamping itu, hasil uji ekotoksikologi terhadap produk Botanigard menunjukkan bahwa risiko secara ekologis yang diperlihatkan oleh serangga bukan sasaran yang diperlakukan dengan formulasi B. bassiana sangat rendah (US EPA, 2006).
Dihubungkan dengan keamanan secara hayati, cendawan entomopatogen dikelompokkan menjadi cendawan dengan kisaran inang spesifik dan yang kisaran inangnya luas (MacLeod, 1963). Cendawan yang memiliki kisaran inang spesifik umumnya menjadi parasit sejati (obligat) dan bersifat sangat virulen tarhadap inang. Sebaliknya yang kisaran inangnya luas sebagian besar merupakan patogen fakultatif, bersifat saprofit, dan cenderung kurang patogenik (Goettel et al., 1990), dan biasanya virulensinya tinggi hanya pada spesies inang dari mana cendawan tersebut pertama kali diisolasi. Contoh, B. bassiana yang diisolasi dari ulat H. armigera akan lebih patogenik pada inangnya tersebut dibanding dengan inang-inangnya yang lain. Selain itu, cendawan yang kisaran inangnya lebih luas justru menjadi lebih spesifik menginfeksi inang jika di lapang. Hal ini dapat terjadi kemungkinan karena dipengaruhi oleh interaksi antara faktor abiotik dan biotik di lapang, sehingga serangga yang mudah terinfeksi di laboratorium belum tentu mudah juga terinfeksi di lapang. Oleh karena itu, kemungkinan terinfeksinya serangga bukan sasaran oleh B. bassiana di lapang sangat kecil. Dengan demikian, aplikasi B. bassiana di lapang cenderung aman bagi musuh alami atau serangga berguna lainnya. Infeksi B. bassiana pada manusia sangat jarang terjadi. Meskipun demikian, dilaporkan ada dua kasus infeksi B. bassiana yang menyebabkan mikosis pada manusia (Henke et al., 2002; Tucker et al., 2004). Namun infeksi tersebut terjadi pada kondisi kesehatan manusia yang sangat buruk akibat penyakit leukimia akut.
Pengujian Botanigard terhadap mamalia, burung, dan ikan juga tidak menunjukkan pengaruh negatif terhadap perkembangan hewan-hewan tersebut (US EPA, 2006). Demikian pula pengujian terhadap sejumlah reptil maupun vertebrata yang membuktikan bahwa B. bassiana tidak menginfeksi keduanya (Georg et al., 1962; Fromtling et al., 1979; Gonzalez et al., 1995).
Cukup banyak tersedia bahan untuk media alami perbanyakan B. bassiana, antara lain: beras, gandum, kedelai, jagung, padi-padian, sorghum, kentang, roti, dan kacang-kacangan. Bahan mana yang akan digunakan tergantung pada beberapa faktor, termasuk kemudahan memperoleh bahan tersebut, biaya, dan strain isolat yang akan diperbanyak. Dalam perbanyakan B. bassiana dengan bahan-bahan alami, untuk menghasilkan konidia dalam jumlah maksimal diperlukan media dengan partikel yang permukaannya lebih luas. Bahan media yang cenderung menggumpal akan memiliki luas permukaan yang sempit, sehingga produksi konidia juga sedikit. Media yang ideal adalah media yang tidak hanya mempunyai partikel dengan permukaan luas, tetapi juga yang dapat mempertahankan keutuhan partikel selama proses produksi (Maheva et al., 1984; Bradley et al., 1992).
Tiga jenis bahan media alami yang telah dicoba dalam perbanyakan B. bassiana skala besar di New Zealand adalah beras, gandum, dan barley. Hasilnya, beras merupakan media paling sesuai bagi perkembangan B. bassiana dengan produktivitas konidia tertinggi mencapai 4,38 x 109 konidia/g beras (Nelson dan Glare, 1996). Penggunaan berbagai jenis sereal, selain beras, sebagai media perbanyakan B. bassiana perlu dipertimbangkan mengingat kandungan nutrisinya yang sangat bervariasi (Jenkins et al., 1998). Perbedaan kandungan nutrisi ini sangat mempengaruhi produksi konidia, terutama per kelompok produksi (batch). Oleh karena itu, pemilihan bahan media perbanyakan harus dilakukan secara tepat, terutama memilih bahan yang memiliki kemampuan produksi konidia secara konsisten dalam kelompok-kelompok produksi. Hasil penelitian lain juga membuktikan bahwa beras putih merupakan bahan media perbanyakan B. bassiana yang tepat karena produksi konidia yang tinggi (Alves dan Pereira, 1989; Mendonca, 1992; Ibrahim dan Low, 1993; Milner et al., 1993). Hal tersebut
Menunjukkan bahwa kombinasi faktor-faktor produkai sangat kompatibel, termasuk keseimbangan nutrisi dalam bahan media, biaya produksi, kemudahan memperoleh bahan, karakter fisik bahan, seperti ukuran, bentuk, dan keutuhan bahan baik sebelum maupun setelah pengkolonisasian konidia.
Dalam perbanyakan, temperatur inkubasi dan cahaya sangat menentukan produktivitas konidia. Temperatur optimal setiap cendawan bervariasi tidak saja antar spesies, tetapi juga antar isolat (Thomas dan Jenkins, 1997; Alasoadura, 1963). Temperatur optimal untuk perkecambahan konidia adalah 25-30°C, dengan temperatur minimum 10°C dan maksimum 32°C. Sedangkan pH optimal untuk pertumbuhan adalah 5,7-5,9, tapi idealnya pH 7-8 (Goral dan Lappa, 1972). Beberapa cendawan membutuhkan cahaya untuk proses sporulasi, sedangkan cendawan lainnya tidak terpengaruh oleh cahaya. Tetapi ada pula cendawan yang sporulasinya terhambat pada tingkat intensitas cahaya tertentu (Vouk dan Klas, 1931). Penelitian terdahulu membuktikan bahwa B. bassiana yang diproduksi di lingkungan tanpa cahaya (gelap) konidianya cenderung berukuran lebih besar dan lebih virulen dibanding yang diproduksi pada tempat terang (Humphreys et al., 1989; Williams, 1959). Hal ini penting sebagai bahan pertimbangan dalam memilih kemasan yang sesuai apabila biakan cendawan harus dibawa ke luar areal perbanyakan. Selain itu yang lebih penting dalam perbanyakan B. bassiana untuk skala komersial adalah kesesuaian produk dengan teknik formulasi dan aplikasinya.
Umumnya produk B. bassiana diformulasi dalam bentuk bubuk (powder) dan merupakan formulasi paling efektif memicu kontak dengan hama sasaran (Stimac et al., 1993). Formulasi B. bassiana berupa pellet hasil enkapsulasi miselium selain efektif untuk meningkatkan mortalitas hama juga untuk mengurangi kompetisi dengan mikroba lain, sehingga meningkatkan daya hidup B. bassiana (White, 1995).
Dalam jurnal diketahui kalau penambahan tepung tapioka sebanyak 1 g dan suhu penyimpanan 5OC, mempunyai potensi yang baik dalam mempertahankan viabilitas spora B. bassiana sekurang-kurangnya sampai dua bulan penyimpanan. Hal tersebut disebabkan oleh kemampuan daya simpan tepung tapioka yang lebih baik daripada tepung beras dan maizena serta didukung oleh suhu rendah yang sesuai untuk mempertahankan viabilitas spora B. bassiana. Terjadi penurunan viabilitas spora kering B. bassiana lebih cepat, seiring dengan semakin meningkatnya dosis pembawa, suhu dan lama penyimpanan. Penyimpanan pada suhu 5OC menunjukkan kemampuan mempertahankan viabilitas spora B. bassiana kering murni lebih lama daripada kondisi suhu 23C dan suhu 29C.
II. PENUTUP
Kesimpulan
Dari pembahasan di atas kita dapat menyimpulkan bahwa:
Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat
Ada beberapa faktor yang mempengaruhi mekanisme infeksi cendawan entomopatogen Beauveria bassiana, faktor yang meliputi diantaranya kelembaban, suhu, iklim, dan tingkat kekebalan tubuh dari inang
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna.
DAFTAR PUSTAKA
Anonim, 2012. Perbanyakan Cendawan Entomopatogen. http: www. Pengendalian%agents-hayati.com.html.diakses pada tanggal 21 maret 2012
Anonim, 2012. Prospek Beauveria bassiana sebagai bioinsectisida. http//:www. Miss Biology Prospek Beauveria bassiana sebagai bioinsectisida.htm. diakses pada tanggal 21 maret 2012
Anonim, 2012. Beauveria bassiana . Wikipedia bahasa Indonesia, ensiklopedia bebas.html. diakses pada tanggal 21 maret 2012
Balai Penelitian Tanaman Hias. 2006. Beauveria bassiana pengendali hama tanaman. Warta Penelitian dan Pengembangan Pertanian Indonesia 28(1): 11-12.
EOL. 2000. Beauveria bassiana Bals. -Criv. Vuill. 1912.[terhubung berkala] http://www.eol.org/pages/160292 . diakses pada tanggal 21 maret 2012
McCoy C, Quintela ED, Faria M de. 1990. Environmental persistence of entomopathogenic fungi. [terhubung berkala]. http://www.lsuagcenter.com/s265/mccoy.htm. diakses pada tanggal 21 maret 2012
cendawan entomopatogen beauveria bassiana
I. PENDAHULUAN
Agens Pengendali Hayati adalah musuh alami berupa Parasitoid, Predator, Entomopatogen dan Agens Antagonis yang telah diketahui mempunyai potensi yang besar sebagai salah satu sarana pengendalian OPT.
Entomopatogen atau penyakit pada serangga hama yang mencakup semua mikroba yaitu Cendawan, bakteri, virus, dan nematoda yang memiliki kemampuan untuk menjadi parasit (pathogen) pada serangga sehingga dapat mematikan serangga yang diinfeksinya. Cendawan adalah mikroba yang mempunyai bentuk tubuh menyerupai benang yang disebut hifa, nampak seperti tumbuhan namun tidak memiliki klorofil.
Mikroba dari jenis jamur adalah Beauveria bassiana, Metarrhizium anisopliae Metarrhizium ftavoviridae Verticillium lecanii dll. dapat mengendalikan Uret, Kepinding tanah, Belalang, Lalat buah dan serangga lainnya.
Kesulitan dan permasalahan utama dalam penerapan dan pengembangan pengendalian hayati adalah modal investasi permulaan yang besar yang harus dikeluarkan untuk kegiatan eksplorasi, penelitian, pengujian dan evaluasi terutama yang menyangkut berbagai aspek dasar baik untuk hama, musuh alami maupun tanaman. Aspek dasar dapat meliputi taksonomi, ekologi, biologi, siklus hidup, dinamika populasi, genetika, fisiologi, dll. Identifikasi yang tepat baik untuk jenis hama maupun musuh alaminya merupakan langkah permulaan yang sangat penting.
Kecuali diperlukan modal, fasilitas yang lengkap juga diperlukan sumber daya manusia terutama para peneliti yang berkualitas dan berpendidikan khusus dan berdedikasi tinggi sesuai dengan yang diperlukan untuk pengembangan teknologi pengendalian hayati. Meskipun ada beberapa ahli yang berpendapat bahwa untuk pengendalian hayati yang penting adalah adanya tenaga peneliti yang berpengalaman dan berdedikasi tinggi serta cukup memiliki rasa seni dan intuisi, namun bagaimanapun untuk keberhasilan pengendalian hayati dalam kerangka PHT diperlukan juga dasar pengetahuan dan teknologi yang mantap.
II. PEMBAHASAN
Adanya kekhawatiran akan pengaruh negatif tentang pemakaian agrokhemikal telah meningkatkan perhatian masyarakat kepada bioinsektisida sebagai alternative teknologi untuk menurunkan populasi hama. Salah satu alternatif pengendalian adalah pemanfaatan jamur penyebab penyakit pada serangga (bioinsectisida), yaitu jamur patogen serangga Beauveria bassiana.
Di Indonesia, hasil-hasil penelitian B. bassiana juga telah banyak dipublikasikan, terutama dari tanaman pangan untuk mengendalikan serangga hama kedelai (Riptortus linearis dan Spodoptera litura), walang sangit pada padi (Leptocoriza acuta) (Prayogo, 2006), Plutella xylostella pada sayursayuran (Hardiyanti, 2006), hama bubuk buah kopi Helopeltis antoni, dan penggerek buah kakaoHypothenemus hampei (Sudarmadji dan Prayogo, dalam Prayogo, 2006).
Saat ini produk bioinsektisida berbahan aktif B. bassiana telah tersedia secara komersial di Indonesia. Meskipun demikian, tampaknya pemanfaatannya di lapang khususnya untuk tanaman perkebunan belum optimal. Padahal,lingkungan mikro tanaman perkebunan sangat ideal bagi perkembangan epizootik cendawan-cendawan entomopatogen, termasuk B. bassiana. Keberlangsungan epizootik cendawan sangat dipengaruhi oleh suhu dan kelembaban lingkungan, dan kriteria ini dapat ditemukan pada tanaman-tanaman perkebunan yang banyak diusahakan di Indonesia. Disamping itu, pemanfaatan cendawan ini dan patogen serangga secara umum dalam pengendalian hama berpotensi memberi keuntungan ekologis jangka panjang terhadap keseimbangan hayati maupun keberlanjutan sistem pertanian.
Pengendalian penyakit tanaman dengan menggunakan agens pengendali hayati muncul karena kekhawatiran masyarakat dunia akibat penggunaan pestisida kimia sintetis. Adanya kekhawatiran tersebut membuat pengendalian hayati menjadi salah satu pilihan cara mengendalikan patogen tanaman yang harus dipertimbangkan.
Agensia hayati adalah setiap organisme yang meliputi spesies, subspesies, varietas, semua jenis serangga, nematoda, protozoa, cendawan (fungi), bakteri, virus, mikoplasma, serta organisme lainnya dalam semua tahap perkembangannya yang dapat dipergunakan untuk keperluan pengendalian hama dan penyakit atau organisme pengganggu, proses produksi, pengolahan hasil pertanian, dan berbagai keperluan lainnya (Permentan No. 411 tahun 1995).
Beauvaria bassiana merupakan cendawan entomopatogen yaitu cendawan yang dapat menimbulkan penyakit pada serangga. Beauveria bassiana secara alami terdapat di dalam tanah sebagai jamur saprofit. Pertumbuhan jamur di dalam tanah sangat dipengaruhi oleh kondisi tanah, seperti kandungan bahan organik, suhu, kelembapan, kebiasaan makan serangga, adanya pestisida sintetis, dan waktu aplikasi. Secara umum, suhu di atas 30 °C, kelembapan tanah yang berkurang dan adanya antifungal atau pestisida dapat menghambat pertumbuhannya.
Beauveria bassiana membunuh hama melalui infeksi sebagai akibat dari serangga yang kontak dengan spora jamur. Serangga dapat kontak dengan spora jamur melalui beberapa cara: semprotan jamur menempel pada tubuh serangga, serangga bergerak pada permukaan tanaman yang sudah terinfeksi jamur, atau dengan memakan jaringan tanaman yang telah diperlakukan dengan jamur. Setelah spora jamur melekat pada kulit serangga (kutikula), mereka berkecambah membentuk struktur (hifa) yang menembus tubuh serangga dan berkembang biak. Proses ini memakan waktu 3-5 hari sampai akhirnya serangga mati, bangkai yang terinfeksi dapat berfungsi sebagai sumber spora untuk penyebaran sekunder jamur. Keberhasilan penyemprotan menggunakan jamur Beauveria bassiana bergantung pada kerentanan spesies yang bersangkutan, tingkat populasi hama, dan kondisi lingkungan pada saat aplikasi, serta sumber daya manusia itu sendiri.
Mekanisme Infeksi Beauveria Bassiana
B. bassiana memproduksi toksin yang disebut beauvericin (Kučera dan Samšiňáková, 1968). Antibiotik ini dapat menyebabkan gangguan pada fungsi hemolimfa dan nukleus serangga, sehingga mengakibatkan pembengkakan yang disertai pengerasan pada serangga yang terinfeksi. Selain secara kontak, B. bassiana juga dapat menginfeksi serangga melalui inokulasi atau kontaminasi pakan. Broome et al. (1976) menyatakan bahwa 37% dari konidia B. bassiana yang dicampurkan ke dalam pakan semut api, Selenopsis richteri, berkecambah di dalam saluran pencernaan inangnya dalam waktu 72 jam, sedangkan hifanya mampu menembus dinding usus antara 60-72 jam. Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat. Serangga yang mati tidak selalu disertai gejala pertumbuhan spora. Contohnya, aphid yang terinfeksi B.bassiana hanya mengalami pembengkakan tanpa terjadi perubahan warna. Demikian pula tempayak lalat yang terinfeksi B. bassiana sering ditemukan secara berkelompok pada ujung-ujung rerumputan (Plate, 1976).
Kematian serangga biasanya disebabkan oleh kerusakan jaringan secara menyeluruh, dan atau karena toksin yang diproduksi oleh cendawan. Menurut Cheung dan Grula (1982), penyakit white muscardine yang menyerang saluran pencernaan Heliothis zea mengakibatkan gangguan nutrisi hingga kematian. Serangga yang terbunuh tubuhnya akan berwarna putih karena ditumbuhi konidia B. bassiana. Jumlah konidia yang dapat dihasilkan oleh satu serangga ditentukan oleh besar kecilnya ukuran serangga tersebut. Setiap serangga terinfeksi B. bassiana akan efektif menjadi sumber infeksi bagi serangga sehat di sekitarnya.
Seperti cendawan lain, perrtumbuhan B. bassiana juga sangat ditentukan oleh kelembapan lingkungan. Namun demikian, cendawan ini juga memiliki fase resisten yang dapat mempertahankan kemampuannya menginfeksi inang pada kondisi kering. Keberadaan epizootiknya di alam menyebabkan B. bassiana secara cepat menginfeksi populasi serangga hingga menyebabkan kematian. Selain itu, kemampuan penetrasinya yang tinggi pada tubuh serangga menyebabkan cendawan ini juga dengan mudah menginfeksi serangga hama pengisap, seperti aphid (Aphis sp.) dan kutu putih Bemisia spp. yang tidak mudah terinfeksi oleh bakteri maupun virus.
Faktor lingkungan, terutama kelembaban dan temperatur serta sedikit cahaya sangat penting perannya dalam proses infeksi dan sporulasi cendawan entomopatogen (Roberts dan Campbell, 1977; McCoy et al., 1988). Temperatur optimum untuk perkembangan, patogenisitas, dan kelulusan hidup cendawan umumnya antara 20-30°C (McCoy et al., 1988). Untuk perkecambahan konidia dan sporulasi pada permukaan tubuh serangga dibutuhkan kelembaban sangat tinggi (> 90% RH), terutama kelembaban di lingkungan mikro sekitar konidia sangat penting perannya dalam proses perkecambahan dan produksi konidia (Millstein et al., 1983; Nordin et al., 1983). Tetapi sebaliknya untuk melepaskan konidia B. bassiana dari konidifor hanya dibutuhkan kelembaban sekitar 50% (Gottwald dan Tedders, 1982).
Meskipun pengaruh cahaya terhadap infeksi cendawan belum diketahui secara jelas, tetapi intensitas sinar ultraviolet tertentu dapat merusak konidia cendawan (Callaghan, 1969). Fuxa (1987) menyatakan bahwa intensitas cahaya matahari dengan rata-rata panjang gelombang antara 290-400 nm cukup efektif menurunkan persistensi deposit konidia pada pertanaman. Sementara Ignoffo et al. (1977) mengemukakan bahwa waktu paruh (half-life) sebagian besar spora cendawan yang terekspos cahaya buatan dengan panjang gelombang mendekati panjang gelombang sinar matahari (290-400 nm) hanya sekitar 1-4 jam, tetapi kenyataannya di lapang waktu paruh dapat mencapai lebih dari 4 jam.
Umumnya cendawan entomopatogen membutuhkan lingkungan yang lembab untuk dapat menginfeksi serangga, oleh karena itu epizootiknya di alam biasanya terbentuk pada saat kondisi lingkungan lembab atau basah. Keefektifan B. bassiana menginfeksi serangga hama tergantung pada spesies atau strain cendawan, dan kepekaan stadia serangga pada tingkat kelembaban lingkungan, struktur tanah (untuk serangga dalam tanah), dan temperatur yang tepat. Selain itu, harus terjadi kontak antara spora B. bassiana yang diterbangkan angin atau terbawa air dengan serangga inang agar terjadi infeksi. Plate (1976) juga menyatakan bahwa epizootik cendawan yang terbentuk secara alami efektif mengendalikan populasi aphid, tempayak lalat yang menyerang perakaran tanaman, belalang, dan thrip, disamping juga potensial sebagai faktor mortalitas utama aphid yang menyerang kentang dan tanaman inang lainnya.
Konidia merupakan unit B. bassiana yang paling infektif dan stabil untuk aplikasi di lapang dibandingkan dengan hifa maupun blastosporanya (Soper dan Ward, 1981; Feng et al., 1994). Konidia yang diaplikasikan dapat berupa suspensi (tidak diformulasi), formulasi butiran, dan bentuk pellet, dan ketiganya memperlihatkan hasil pengendalian yang cukup nyata. Stimac et al. (1993) menyatakan bahwa aplikasi konidia B. bassiana dengan cara sprinkle dan disemprotkan pada permukaan tanah sangat efektif menyebabkan mortalitas hama sasaran. Mortalitas hama semut api, Selenopsis invicta, yang dikendalikan dengan B. bassiana tertinggi mulai 3-8 hari setelah perlakuan. Sedangkan enkapsulasi (pellet) konidia B. bassiana dengan menggunakan kalsium alginat juga efektif meningkatkan mortalitas S. invicta (White, 1995), karena enkapsulasi menyebabkan konidia lebih stabil di dalam tanah.
Beberapa senyawa metabolit sekunder diproduksi oleh B. bassiana, seperti beauvericin, bassianin, bassiacridin, bassianolide, beauverolides, tenellin, dan oosporein (Strasser et al., 2000; Vey et al., 2001; Quesada-Moraga dan Vey, 2004). Senyawa metabolit sekunder ini dapat dihasilkan oleh B. bassiana pada epizootik di alam (tanah) maupun pada epizootik buatan (di laboratorium). Meskipun demikian, hingga saat ini belum ada laporan tentang tercemarnya rantai makanan oleh senyawa metabolit sekunder, atau terakumulasi di alam sebagai limbah epizootik B. bassiana (Vey et al., 2001).
Penggunaan B. bassiana dalam pengendalian hama telah diuji secara luas di berbagai negara. Hasil uji toksikologi terhadap salah satu produk B. bassiana, Botanigard, menunjukkan bahwa produk tersebut tidak menimbulkan dampak negatif yang berhubungan dengan patogenisitas dan toksisitasnya, sehingga produk tersebut digunakan secara aman selama lebih dari 10 tahun di Amerika Serikat dan juga di beberapa negara lain (US EPA, 2006).
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna. Namun, Plate (1976) mengungkapkan bahwa tingkat kepekaan serangga bukan sasaran terhadap infeksi B. bassiana sangat ditentukan oleh virulensi dan patogenisitas cendawan, serta spesies serangga inang. Selain itu, perbedaan fisiologis dan ekologis inang juga mempengaruhi infeksi B. bassiana. Misalnya, serangga bukan sasaran yang mudah terinfeksi B. bassiana di laboratorium tidak akan serta merta terinfeksi pada kondisi lapang. Ludwig dan Oetting (2001) menegaskan bahwa beberapa serangga musuh alami yang peka terhadap infeksi B. bassiana di laboratorium ternyata mengalami infeksi sangat rendah pada uji di rumah kaca. Disamping itu, hasil uji ekotoksikologi terhadap produk Botanigard menunjukkan bahwa risiko secara ekologis yang diperlihatkan oleh serangga bukan sasaran yang diperlakukan dengan formulasi B. bassiana sangat rendah (US EPA, 2006).
Dihubungkan dengan keamanan secara hayati, cendawan entomopatogen dikelompokkan menjadi cendawan dengan kisaran inang spesifik dan yang kisaran inangnya luas (MacLeod, 1963). Cendawan yang memiliki kisaran inang spesifik umumnya menjadi parasit sejati (obligat) dan bersifat sangat virulen tarhadap inang. Sebaliknya yang kisaran inangnya luas sebagian besar merupakan patogen fakultatif, bersifat saprofit, dan cenderung kurang patogenik (Goettel et al., 1990), dan biasanya virulensinya tinggi hanya pada spesies inang dari mana cendawan tersebut pertama kali diisolasi. Contoh, B. bassiana yang diisolasi dari ulat H. armigera akan lebih patogenik pada inangnya tersebut dibanding dengan inang-inangnya yang lain. Selain itu, cendawan yang kisaran inangnya lebih luas justru menjadi lebih spesifik menginfeksi inang jika di lapang. Hal ini dapat terjadi kemungkinan karena dipengaruhi oleh interaksi antara faktor abiotik dan biotik di lapang, sehingga serangga yang mudah terinfeksi di laboratorium belum tentu mudah juga terinfeksi di lapang. Oleh karena itu, kemungkinan terinfeksinya serangga bukan sasaran oleh B. bassiana di lapang sangat kecil. Dengan demikian, aplikasi B. bassiana di lapang cenderung aman bagi musuh alami atau serangga berguna lainnya. Infeksi B. bassiana pada manusia sangat jarang terjadi. Meskipun demikian, dilaporkan ada dua kasus infeksi B. bassiana yang menyebabkan mikosis pada manusia (Henke et al., 2002; Tucker et al., 2004). Namun infeksi tersebut terjadi pada kondisi kesehatan manusia yang sangat buruk akibat penyakit leukimia akut.
Pengujian Botanigard terhadap mamalia, burung, dan ikan juga tidak menunjukkan pengaruh negatif terhadap perkembangan hewan-hewan tersebut (US EPA, 2006). Demikian pula pengujian terhadap sejumlah reptil maupun vertebrata yang membuktikan bahwa B. bassiana tidak menginfeksi keduanya (Georg et al., 1962; Fromtling et al., 1979; Gonzalez et al., 1995).
Cukup banyak tersedia bahan untuk media alami perbanyakan B. bassiana, antara lain: beras, gandum, kedelai, jagung, padi-padian, sorghum, kentang, roti, dan kacang-kacangan. Bahan mana yang akan digunakan tergantung pada beberapa faktor, termasuk kemudahan memperoleh bahan tersebut, biaya, dan strain isolat yang akan diperbanyak. Dalam perbanyakan B. bassiana dengan bahan-bahan alami, untuk menghasilkan konidia dalam jumlah maksimal diperlukan media dengan partikel yang permukaannya lebih luas. Bahan media yang cenderung menggumpal akan memiliki luas permukaan yang sempit, sehingga produksi konidia juga sedikit. Media yang ideal adalah media yang tidak hanya mempunyai partikel dengan permukaan luas, tetapi juga yang dapat mempertahankan keutuhan partikel selama proses produksi (Maheva et al., 1984; Bradley et al., 1992).
Tiga jenis bahan media alami yang telah dicoba dalam perbanyakan B. bassiana skala besar di New Zealand adalah beras, gandum, dan barley. Hasilnya, beras merupakan media paling sesuai bagi perkembangan B. bassiana dengan produktivitas konidia tertinggi mencapai 4,38 x 109 konidia/g beras (Nelson dan Glare, 1996). Penggunaan berbagai jenis sereal, selain beras, sebagai media perbanyakan B. bassiana perlu dipertimbangkan mengingat kandungan nutrisinya yang sangat bervariasi (Jenkins et al., 1998). Perbedaan kandungan nutrisi ini sangat mempengaruhi produksi konidia, terutama per kelompok produksi (batch). Oleh karena itu, pemilihan bahan media perbanyakan harus dilakukan secara tepat, terutama memilih bahan yang memiliki kemampuan produksi konidia secara konsisten dalam kelompok-kelompok produksi. Hasil penelitian lain juga membuktikan bahwa beras putih merupakan bahan media perbanyakan B. bassiana yang tepat karena produksi konidia yang tinggi (Alves dan Pereira, 1989; Mendonca, 1992; Ibrahim dan Low, 1993; Milner et al., 1993). Hal tersebut
Menunjukkan bahwa kombinasi faktor-faktor produkai sangat kompatibel, termasuk keseimbangan nutrisi dalam bahan media, biaya produksi, kemudahan memperoleh bahan, karakter fisik bahan, seperti ukuran, bentuk, dan keutuhan bahan baik sebelum maupun setelah pengkolonisasian konidia.
Dalam perbanyakan, temperatur inkubasi dan cahaya sangat menentukan produktivitas konidia. Temperatur optimal setiap cendawan bervariasi tidak saja antar spesies, tetapi juga antar isolat (Thomas dan Jenkins, 1997; Alasoadura, 1963). Temperatur optimal untuk perkecambahan konidia adalah 25-30°C, dengan temperatur minimum 10°C dan maksimum 32°C. Sedangkan pH optimal untuk pertumbuhan adalah 5,7-5,9, tapi idealnya pH 7-8 (Goral dan Lappa, 1972). Beberapa cendawan membutuhkan cahaya untuk proses sporulasi, sedangkan cendawan lainnya tidak terpengaruh oleh cahaya. Tetapi ada pula cendawan yang sporulasinya terhambat pada tingkat intensitas cahaya tertentu (Vouk dan Klas, 1931). Penelitian terdahulu membuktikan bahwa B. bassiana yang diproduksi di lingkungan tanpa cahaya (gelap) konidianya cenderung berukuran lebih besar dan lebih virulen dibanding yang diproduksi pada tempat terang (Humphreys et al., 1989; Williams, 1959). Hal ini penting sebagai bahan pertimbangan dalam memilih kemasan yang sesuai apabila biakan cendawan harus dibawa ke luar areal perbanyakan. Selain itu yang lebih penting dalam perbanyakan B. bassiana untuk skala komersial adalah kesesuaian produk dengan teknik formulasi dan aplikasinya.
Umumnya produk B. bassiana diformulasi dalam bentuk bubuk (powder) dan merupakan formulasi paling efektif memicu kontak dengan hama sasaran (Stimac et al., 1993). Formulasi B. bassiana berupa pellet hasil enkapsulasi miselium selain efektif untuk meningkatkan mortalitas hama juga untuk mengurangi kompetisi dengan mikroba lain, sehingga meningkatkan daya hidup B. bassiana (White, 1995).
Dalam jurnal diketahui kalau penambahan tepung tapioka sebanyak 1 g dan suhu penyimpanan 5OC, mempunyai potensi yang baik dalam mempertahankan viabilitas spora B. bassiana sekurang-kurangnya sampai dua bulan penyimpanan. Hal tersebut disebabkan oleh kemampuan daya simpan tepung tapioka yang lebih baik daripada tepung beras dan maizena serta didukung oleh suhu rendah yang sesuai untuk mempertahankan viabilitas spora B. bassiana. Terjadi penurunan viabilitas spora kering B. bassiana lebih cepat, seiring dengan semakin meningkatnya dosis pembawa, suhu dan lama penyimpanan. Penyimpanan pada suhu 5OC menunjukkan kemampuan mempertahankan viabilitas spora B. bassiana kering murni lebih lama daripada kondisi suhu 23C dan suhu 29C.
II. PENUTUP
Kesimpulan
Dari pembahasan di atas kita dapat menyimpulkan bahwa:
Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat
Ada beberapa faktor yang mempengaruhi mekanisme infeksi cendawan entomopatogen Beauveria bassiana, faktor yang meliputi diantaranya kelembaban, suhu, iklim, dan tingkat kekebalan tubuh dari inang
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna.
DAFTAR PUSTAKA
Anonim, 2012. Perbanyakan Cendawan Entomopatogen. http: www. Pengendalian%agents-hayati.com.html.diakses pada tanggal 21 maret 2012
Anonim, 2012. Prospek Beauveria bassiana sebagai bioinsectisida. http//:www. Miss Biology Prospek Beauveria bassiana sebagai bioinsectisida.htm. diakses pada tanggal 21 maret 2012
Anonim, 2012. Beauveria bassiana . Wikipedia bahasa Indonesia, ensiklopedia bebas.html. diakses pada tanggal 21 maret 2012
Balai Penelitian Tanaman Hias. 2006. Beauveria bassiana pengendali hama tanaman. Warta Penelitian dan Pengembangan Pertanian Indonesia 28(1): 11-12.
EOL. 2000. Beauveria bassiana Bals. -Criv. Vuill. 1912.[terhubung berkala] http://www.eol.org/pages/160292 . diakses pada tanggal 21 maret 2012
McCoy C, Quintela ED, Faria M de. 1990. Environmental persistence of entomopathogenic fungi. [terhubung berkala]. http://www.lsuagcenter.com/s265/mccoy.htm. diakses pada tanggal 21 maret 2012
I. PENDAHULUAN
Agens Pengendali Hayati adalah musuh alami berupa Parasitoid, Predator, Entomopatogen dan Agens Antagonis yang telah diketahui mempunyai potensi yang besar sebagai salah satu sarana pengendalian OPT.
Entomopatogen atau penyakit pada serangga hama yang mencakup semua mikroba yaitu Cendawan, bakteri, virus, dan nematoda yang memiliki kemampuan untuk menjadi parasit (pathogen) pada serangga sehingga dapat mematikan serangga yang diinfeksinya. Cendawan adalah mikroba yang mempunyai bentuk tubuh menyerupai benang yang disebut hifa, nampak seperti tumbuhan namun tidak memiliki klorofil.
Mikroba dari jenis jamur adalah Beauveria bassiana, Metarrhizium anisopliae Metarrhizium ftavoviridae Verticillium lecanii dll. dapat mengendalikan Uret, Kepinding tanah, Belalang, Lalat buah dan serangga lainnya.
Kesulitan dan permasalahan utama dalam penerapan dan pengembangan pengendalian hayati adalah modal investasi permulaan yang besar yang harus dikeluarkan untuk kegiatan eksplorasi, penelitian, pengujian dan evaluasi terutama yang menyangkut berbagai aspek dasar baik untuk hama, musuh alami maupun tanaman. Aspek dasar dapat meliputi taksonomi, ekologi, biologi, siklus hidup, dinamika populasi, genetika, fisiologi, dll. Identifikasi yang tepat baik untuk jenis hama maupun musuh alaminya merupakan langkah permulaan yang sangat penting.
Kecuali diperlukan modal, fasilitas yang lengkap juga diperlukan sumber daya manusia terutama para peneliti yang berkualitas dan berpendidikan khusus dan berdedikasi tinggi sesuai dengan yang diperlukan untuk pengembangan teknologi pengendalian hayati. Meskipun ada beberapa ahli yang berpendapat bahwa untuk pengendalian hayati yang penting adalah adanya tenaga peneliti yang berpengalaman dan berdedikasi tinggi serta cukup memiliki rasa seni dan intuisi, namun bagaimanapun untuk keberhasilan pengendalian hayati dalam kerangka PHT diperlukan juga dasar pengetahuan dan teknologi yang mantap.
II. PEMBAHASAN
Adanya kekhawatiran akan pengaruh negatif tentang pemakaian agrokhemikal telah meningkatkan perhatian masyarakat kepada bioinsektisida sebagai alternative teknologi untuk menurunkan populasi hama. Salah satu alternatif pengendalian adalah pemanfaatan jamur penyebab penyakit pada serangga (bioinsectisida), yaitu jamur patogen serangga Beauveria bassiana.
Di Indonesia, hasil-hasil penelitian B. bassiana juga telah banyak dipublikasikan, terutama dari tanaman pangan untuk mengendalikan serangga hama kedelai (Riptortus linearis dan Spodoptera litura), walang sangit pada padi (Leptocoriza acuta) (Prayogo, 2006), Plutella xylostella pada sayursayuran (Hardiyanti, 2006), hama bubuk buah kopi Helopeltis antoni, dan penggerek buah kakaoHypothenemus hampei (Sudarmadji dan Prayogo, dalam Prayogo, 2006).
Saat ini produk bioinsektisida berbahan aktif B. bassiana telah tersedia secara komersial di Indonesia. Meskipun demikian, tampaknya pemanfaatannya di lapang khususnya untuk tanaman perkebunan belum optimal. Padahal,lingkungan mikro tanaman perkebunan sangat ideal bagi perkembangan epizootik cendawan-cendawan entomopatogen, termasuk B. bassiana. Keberlangsungan epizootik cendawan sangat dipengaruhi oleh suhu dan kelembaban lingkungan, dan kriteria ini dapat ditemukan pada tanaman-tanaman perkebunan yang banyak diusahakan di Indonesia. Disamping itu, pemanfaatan cendawan ini dan patogen serangga secara umum dalam pengendalian hama berpotensi memberi keuntungan ekologis jangka panjang terhadap keseimbangan hayati maupun keberlanjutan sistem pertanian.
Pengendalian penyakit tanaman dengan menggunakan agens pengendali hayati muncul karena kekhawatiran masyarakat dunia akibat penggunaan pestisida kimia sintetis. Adanya kekhawatiran tersebut membuat pengendalian hayati menjadi salah satu pilihan cara mengendalikan patogen tanaman yang harus dipertimbangkan.
Agensia hayati adalah setiap organisme yang meliputi spesies, subspesies, varietas, semua jenis serangga, nematoda, protozoa, cendawan (fungi), bakteri, virus, mikoplasma, serta organisme lainnya dalam semua tahap perkembangannya yang dapat dipergunakan untuk keperluan pengendalian hama dan penyakit atau organisme pengganggu, proses produksi, pengolahan hasil pertanian, dan berbagai keperluan lainnya (Permentan No. 411 tahun 1995).
Beauvaria bassiana merupakan cendawan entomopatogen yaitu cendawan yang dapat menimbulkan penyakit pada serangga. Beauveria bassiana secara alami terdapat di dalam tanah sebagai jamur saprofit. Pertumbuhan jamur di dalam tanah sangat dipengaruhi oleh kondisi tanah, seperti kandungan bahan organik, suhu, kelembapan, kebiasaan makan serangga, adanya pestisida sintetis, dan waktu aplikasi. Secara umum, suhu di atas 30 °C, kelembapan tanah yang berkurang dan adanya antifungal atau pestisida dapat menghambat pertumbuhannya.
Beauveria bassiana membunuh hama melalui infeksi sebagai akibat dari serangga yang kontak dengan spora jamur. Serangga dapat kontak dengan spora jamur melalui beberapa cara: semprotan jamur menempel pada tubuh serangga, serangga bergerak pada permukaan tanaman yang sudah terinfeksi jamur, atau dengan memakan jaringan tanaman yang telah diperlakukan dengan jamur. Setelah spora jamur melekat pada kulit serangga (kutikula), mereka berkecambah membentuk struktur (hifa) yang menembus tubuh serangga dan berkembang biak. Proses ini memakan waktu 3-5 hari sampai akhirnya serangga mati, bangkai yang terinfeksi dapat berfungsi sebagai sumber spora untuk penyebaran sekunder jamur. Keberhasilan penyemprotan menggunakan jamur Beauveria bassiana bergantung pada kerentanan spesies yang bersangkutan, tingkat populasi hama, dan kondisi lingkungan pada saat aplikasi, serta sumber daya manusia itu sendiri.
Mekanisme Infeksi Beauveria Bassiana
B. bassiana memproduksi toksin yang disebut beauvericin (Kučera dan Samšiňáková, 1968). Antibiotik ini dapat menyebabkan gangguan pada fungsi hemolimfa dan nukleus serangga, sehingga mengakibatkan pembengkakan yang disertai pengerasan pada serangga yang terinfeksi. Selain secara kontak, B. bassiana juga dapat menginfeksi serangga melalui inokulasi atau kontaminasi pakan. Broome et al. (1976) menyatakan bahwa 37% dari konidia B. bassiana yang dicampurkan ke dalam pakan semut api, Selenopsis richteri, berkecambah di dalam saluran pencernaan inangnya dalam waktu 72 jam, sedangkan hifanya mampu menembus dinding usus antara 60-72 jam. Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat. Serangga yang mati tidak selalu disertai gejala pertumbuhan spora. Contohnya, aphid yang terinfeksi B.bassiana hanya mengalami pembengkakan tanpa terjadi perubahan warna. Demikian pula tempayak lalat yang terinfeksi B. bassiana sering ditemukan secara berkelompok pada ujung-ujung rerumputan (Plate, 1976).
Kematian serangga biasanya disebabkan oleh kerusakan jaringan secara menyeluruh, dan atau karena toksin yang diproduksi oleh cendawan. Menurut Cheung dan Grula (1982), penyakit white muscardine yang menyerang saluran pencernaan Heliothis zea mengakibatkan gangguan nutrisi hingga kematian. Serangga yang terbunuh tubuhnya akan berwarna putih karena ditumbuhi konidia B. bassiana. Jumlah konidia yang dapat dihasilkan oleh satu serangga ditentukan oleh besar kecilnya ukuran serangga tersebut. Setiap serangga terinfeksi B. bassiana akan efektif menjadi sumber infeksi bagi serangga sehat di sekitarnya.
Seperti cendawan lain, perrtumbuhan B. bassiana juga sangat ditentukan oleh kelembapan lingkungan. Namun demikian, cendawan ini juga memiliki fase resisten yang dapat mempertahankan kemampuannya menginfeksi inang pada kondisi kering. Keberadaan epizootiknya di alam menyebabkan B. bassiana secara cepat menginfeksi populasi serangga hingga menyebabkan kematian. Selain itu, kemampuan penetrasinya yang tinggi pada tubuh serangga menyebabkan cendawan ini juga dengan mudah menginfeksi serangga hama pengisap, seperti aphid (Aphis sp.) dan kutu putih Bemisia spp. yang tidak mudah terinfeksi oleh bakteri maupun virus.
Faktor lingkungan, terutama kelembaban dan temperatur serta sedikit cahaya sangat penting perannya dalam proses infeksi dan sporulasi cendawan entomopatogen (Roberts dan Campbell, 1977; McCoy et al., 1988). Temperatur optimum untuk perkembangan, patogenisitas, dan kelulusan hidup cendawan umumnya antara 20-30°C (McCoy et al., 1988). Untuk perkecambahan konidia dan sporulasi pada permukaan tubuh serangga dibutuhkan kelembaban sangat tinggi (> 90% RH), terutama kelembaban di lingkungan mikro sekitar konidia sangat penting perannya dalam proses perkecambahan dan produksi konidia (Millstein et al., 1983; Nordin et al., 1983). Tetapi sebaliknya untuk melepaskan konidia B. bassiana dari konidifor hanya dibutuhkan kelembaban sekitar 50% (Gottwald dan Tedders, 1982).
Meskipun pengaruh cahaya terhadap infeksi cendawan belum diketahui secara jelas, tetapi intensitas sinar ultraviolet tertentu dapat merusak konidia cendawan (Callaghan, 1969). Fuxa (1987) menyatakan bahwa intensitas cahaya matahari dengan rata-rata panjang gelombang antara 290-400 nm cukup efektif menurunkan persistensi deposit konidia pada pertanaman. Sementara Ignoffo et al. (1977) mengemukakan bahwa waktu paruh (half-life) sebagian besar spora cendawan yang terekspos cahaya buatan dengan panjang gelombang mendekati panjang gelombang sinar matahari (290-400 nm) hanya sekitar 1-4 jam, tetapi kenyataannya di lapang waktu paruh dapat mencapai lebih dari 4 jam.
Umumnya cendawan entomopatogen membutuhkan lingkungan yang lembab untuk dapat menginfeksi serangga, oleh karena itu epizootiknya di alam biasanya terbentuk pada saat kondisi lingkungan lembab atau basah. Keefektifan B. bassiana menginfeksi serangga hama tergantung pada spesies atau strain cendawan, dan kepekaan stadia serangga pada tingkat kelembaban lingkungan, struktur tanah (untuk serangga dalam tanah), dan temperatur yang tepat. Selain itu, harus terjadi kontak antara spora B. bassiana yang diterbangkan angin atau terbawa air dengan serangga inang agar terjadi infeksi. Plate (1976) juga menyatakan bahwa epizootik cendawan yang terbentuk secara alami efektif mengendalikan populasi aphid, tempayak lalat yang menyerang perakaran tanaman, belalang, dan thrip, disamping juga potensial sebagai faktor mortalitas utama aphid yang menyerang kentang dan tanaman inang lainnya.
Konidia merupakan unit B. bassiana yang paling infektif dan stabil untuk aplikasi di lapang dibandingkan dengan hifa maupun blastosporanya (Soper dan Ward, 1981; Feng et al., 1994). Konidia yang diaplikasikan dapat berupa suspensi (tidak diformulasi), formulasi butiran, dan bentuk pellet, dan ketiganya memperlihatkan hasil pengendalian yang cukup nyata. Stimac et al. (1993) menyatakan bahwa aplikasi konidia B. bassiana dengan cara sprinkle dan disemprotkan pada permukaan tanah sangat efektif menyebabkan mortalitas hama sasaran. Mortalitas hama semut api, Selenopsis invicta, yang dikendalikan dengan B. bassiana tertinggi mulai 3-8 hari setelah perlakuan. Sedangkan enkapsulasi (pellet) konidia B. bassiana dengan menggunakan kalsium alginat juga efektif meningkatkan mortalitas S. invicta (White, 1995), karena enkapsulasi menyebabkan konidia lebih stabil di dalam tanah.
Beberapa senyawa metabolit sekunder diproduksi oleh B. bassiana, seperti beauvericin, bassianin, bassiacridin, bassianolide, beauverolides, tenellin, dan oosporein (Strasser et al., 2000; Vey et al., 2001; Quesada-Moraga dan Vey, 2004). Senyawa metabolit sekunder ini dapat dihasilkan oleh B. bassiana pada epizootik di alam (tanah) maupun pada epizootik buatan (di laboratorium). Meskipun demikian, hingga saat ini belum ada laporan tentang tercemarnya rantai makanan oleh senyawa metabolit sekunder, atau terakumulasi di alam sebagai limbah epizootik B. bassiana (Vey et al., 2001).
Penggunaan B. bassiana dalam pengendalian hama telah diuji secara luas di berbagai negara. Hasil uji toksikologi terhadap salah satu produk B. bassiana, Botanigard, menunjukkan bahwa produk tersebut tidak menimbulkan dampak negatif yang berhubungan dengan patogenisitas dan toksisitasnya, sehingga produk tersebut digunakan secara aman selama lebih dari 10 tahun di Amerika Serikat dan juga di beberapa negara lain (US EPA, 2006).
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna. Namun, Plate (1976) mengungkapkan bahwa tingkat kepekaan serangga bukan sasaran terhadap infeksi B. bassiana sangat ditentukan oleh virulensi dan patogenisitas cendawan, serta spesies serangga inang. Selain itu, perbedaan fisiologis dan ekologis inang juga mempengaruhi infeksi B. bassiana. Misalnya, serangga bukan sasaran yang mudah terinfeksi B. bassiana di laboratorium tidak akan serta merta terinfeksi pada kondisi lapang. Ludwig dan Oetting (2001) menegaskan bahwa beberapa serangga musuh alami yang peka terhadap infeksi B. bassiana di laboratorium ternyata mengalami infeksi sangat rendah pada uji di rumah kaca. Disamping itu, hasil uji ekotoksikologi terhadap produk Botanigard menunjukkan bahwa risiko secara ekologis yang diperlihatkan oleh serangga bukan sasaran yang diperlakukan dengan formulasi B. bassiana sangat rendah (US EPA, 2006).
Dihubungkan dengan keamanan secara hayati, cendawan entomopatogen dikelompokkan menjadi cendawan dengan kisaran inang spesifik dan yang kisaran inangnya luas (MacLeod, 1963). Cendawan yang memiliki kisaran inang spesifik umumnya menjadi parasit sejati (obligat) dan bersifat sangat virulen tarhadap inang. Sebaliknya yang kisaran inangnya luas sebagian besar merupakan patogen fakultatif, bersifat saprofit, dan cenderung kurang patogenik (Goettel et al., 1990), dan biasanya virulensinya tinggi hanya pada spesies inang dari mana cendawan tersebut pertama kali diisolasi. Contoh, B. bassiana yang diisolasi dari ulat H. armigera akan lebih patogenik pada inangnya tersebut dibanding dengan inang-inangnya yang lain. Selain itu, cendawan yang kisaran inangnya lebih luas justru menjadi lebih spesifik menginfeksi inang jika di lapang. Hal ini dapat terjadi kemungkinan karena dipengaruhi oleh interaksi antara faktor abiotik dan biotik di lapang, sehingga serangga yang mudah terinfeksi di laboratorium belum tentu mudah juga terinfeksi di lapang. Oleh karena itu, kemungkinan terinfeksinya serangga bukan sasaran oleh B. bassiana di lapang sangat kecil. Dengan demikian, aplikasi B. bassiana di lapang cenderung aman bagi musuh alami atau serangga berguna lainnya. Infeksi B. bassiana pada manusia sangat jarang terjadi. Meskipun demikian, dilaporkan ada dua kasus infeksi B. bassiana yang menyebabkan mikosis pada manusia (Henke et al., 2002; Tucker et al., 2004). Namun infeksi tersebut terjadi pada kondisi kesehatan manusia yang sangat buruk akibat penyakit leukimia akut.
Pengujian Botanigard terhadap mamalia, burung, dan ikan juga tidak menunjukkan pengaruh negatif terhadap perkembangan hewan-hewan tersebut (US EPA, 2006). Demikian pula pengujian terhadap sejumlah reptil maupun vertebrata yang membuktikan bahwa B. bassiana tidak menginfeksi keduanya (Georg et al., 1962; Fromtling et al., 1979; Gonzalez et al., 1995).
Cukup banyak tersedia bahan untuk media alami perbanyakan B. bassiana, antara lain: beras, gandum, kedelai, jagung, padi-padian, sorghum, kentang, roti, dan kacang-kacangan. Bahan mana yang akan digunakan tergantung pada beberapa faktor, termasuk kemudahan memperoleh bahan tersebut, biaya, dan strain isolat yang akan diperbanyak. Dalam perbanyakan B. bassiana dengan bahan-bahan alami, untuk menghasilkan konidia dalam jumlah maksimal diperlukan media dengan partikel yang permukaannya lebih luas. Bahan media yang cenderung menggumpal akan memiliki luas permukaan yang sempit, sehingga produksi konidia juga sedikit. Media yang ideal adalah media yang tidak hanya mempunyai partikel dengan permukaan luas, tetapi juga yang dapat mempertahankan keutuhan partikel selama proses produksi (Maheva et al., 1984; Bradley et al., 1992).
Tiga jenis bahan media alami yang telah dicoba dalam perbanyakan B. bassiana skala besar di New Zealand adalah beras, gandum, dan barley. Hasilnya, beras merupakan media paling sesuai bagi perkembangan B. bassiana dengan produktivitas konidia tertinggi mencapai 4,38 x 109 konidia/g beras (Nelson dan Glare, 1996). Penggunaan berbagai jenis sereal, selain beras, sebagai media perbanyakan B. bassiana perlu dipertimbangkan mengingat kandungan nutrisinya yang sangat bervariasi (Jenkins et al., 1998). Perbedaan kandungan nutrisi ini sangat mempengaruhi produksi konidia, terutama per kelompok produksi (batch). Oleh karena itu, pemilihan bahan media perbanyakan harus dilakukan secara tepat, terutama memilih bahan yang memiliki kemampuan produksi konidia secara konsisten dalam kelompok-kelompok produksi. Hasil penelitian lain juga membuktikan bahwa beras putih merupakan bahan media perbanyakan B. bassiana yang tepat karena produksi konidia yang tinggi (Alves dan Pereira, 1989; Mendonca, 1992; Ibrahim dan Low, 1993; Milner et al., 1993). Hal tersebut
Menunjukkan bahwa kombinasi faktor-faktor produkai sangat kompatibel, termasuk keseimbangan nutrisi dalam bahan media, biaya produksi, kemudahan memperoleh bahan, karakter fisik bahan, seperti ukuran, bentuk, dan keutuhan bahan baik sebelum maupun setelah pengkolonisasian konidia.
Dalam perbanyakan, temperatur inkubasi dan cahaya sangat menentukan produktivitas konidia. Temperatur optimal setiap cendawan bervariasi tidak saja antar spesies, tetapi juga antar isolat (Thomas dan Jenkins, 1997; Alasoadura, 1963). Temperatur optimal untuk perkecambahan konidia adalah 25-30°C, dengan temperatur minimum 10°C dan maksimum 32°C. Sedangkan pH optimal untuk pertumbuhan adalah 5,7-5,9, tapi idealnya pH 7-8 (Goral dan Lappa, 1972). Beberapa cendawan membutuhkan cahaya untuk proses sporulasi, sedangkan cendawan lainnya tidak terpengaruh oleh cahaya. Tetapi ada pula cendawan yang sporulasinya terhambat pada tingkat intensitas cahaya tertentu (Vouk dan Klas, 1931). Penelitian terdahulu membuktikan bahwa B. bassiana yang diproduksi di lingkungan tanpa cahaya (gelap) konidianya cenderung berukuran lebih besar dan lebih virulen dibanding yang diproduksi pada tempat terang (Humphreys et al., 1989; Williams, 1959). Hal ini penting sebagai bahan pertimbangan dalam memilih kemasan yang sesuai apabila biakan cendawan harus dibawa ke luar areal perbanyakan. Selain itu yang lebih penting dalam perbanyakan B. bassiana untuk skala komersial adalah kesesuaian produk dengan teknik formulasi dan aplikasinya.
Umumnya produk B. bassiana diformulasi dalam bentuk bubuk (powder) dan merupakan formulasi paling efektif memicu kontak dengan hama sasaran (Stimac et al., 1993). Formulasi B. bassiana berupa pellet hasil enkapsulasi miselium selain efektif untuk meningkatkan mortalitas hama juga untuk mengurangi kompetisi dengan mikroba lain, sehingga meningkatkan daya hidup B. bassiana (White, 1995).
Dalam jurnal diketahui kalau penambahan tepung tapioka sebanyak 1 g dan suhu penyimpanan 5OC, mempunyai potensi yang baik dalam mempertahankan viabilitas spora B. bassiana sekurang-kurangnya sampai dua bulan penyimpanan. Hal tersebut disebabkan oleh kemampuan daya simpan tepung tapioka yang lebih baik daripada tepung beras dan maizena serta didukung oleh suhu rendah yang sesuai untuk mempertahankan viabilitas spora B. bassiana. Terjadi penurunan viabilitas spora kering B. bassiana lebih cepat, seiring dengan semakin meningkatnya dosis pembawa, suhu dan lama penyimpanan. Penyimpanan pada suhu 5OC menunjukkan kemampuan mempertahankan viabilitas spora B. bassiana kering murni lebih lama daripada kondisi suhu 23C dan suhu 29C.
II. PENUTUP
Kesimpulan
Dari pembahasan di atas kita dapat menyimpulkan bahwa:
Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat
Ada beberapa faktor yang mempengaruhi mekanisme infeksi cendawan entomopatogen Beauveria bassiana, faktor yang meliputi diantaranya kelembaban, suhu, iklim, dan tingkat kekebalan tubuh dari inang
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna.
DAFTAR PUSTAKA
Anonim, 2012. Perbanyakan Cendawan Entomopatogen. http: www. Pengendalian%agents-hayati.com.html.diakses pada tanggal 21 maret 2012
Anonim, 2012. Prospek Beauveria bassiana sebagai bioinsectisida. http//:www. Miss Biology Prospek Beauveria bassiana sebagai bioinsectisida.htm. diakses pada tanggal 21 maret 2012
Anonim, 2012. Beauveria bassiana . Wikipedia bahasa Indonesia, ensiklopedia bebas.html. diakses pada tanggal 21 maret 2012
Balai Penelitian Tanaman Hias. 2006. Beauveria bassiana pengendali hama tanaman. Warta Penelitian dan Pengembangan Pertanian Indonesia 28(1): 11-12.
EOL. 2000. Beauveria bassiana Bals. -Criv. Vuill. 1912.[terhubung berkala] http://www.eol.org/pages/160292 . diakses pada tanggal 21 maret 2012
McCoy C, Quintela ED, Faria M de. 1990. Environmental persistence of entomopathogenic fungi. [terhubung berkala]. http://www.lsuagcenter.com/s265/mccoy.htm. diakses pada tanggal 21 maret 2012
Agens Pengendali Hayati adalah musuh alami berupa Parasitoid, Predator, Entomopatogen dan Agens Antagonis yang telah diketahui mempunyai potensi yang besar sebagai salah satu sarana pengendalian OPT.
Entomopatogen atau penyakit pada serangga hama yang mencakup semua mikroba yaitu Cendawan, bakteri, virus, dan nematoda yang memiliki kemampuan untuk menjadi parasit (pathogen) pada serangga sehingga dapat mematikan serangga yang diinfeksinya. Cendawan adalah mikroba yang mempunyai bentuk tubuh menyerupai benang yang disebut hifa, nampak seperti tumbuhan namun tidak memiliki klorofil.
Mikroba dari jenis jamur adalah Beauveria bassiana, Metarrhizium anisopliae Metarrhizium ftavoviridae Verticillium lecanii dll. dapat mengendalikan Uret, Kepinding tanah, Belalang, Lalat buah dan serangga lainnya.
Kesulitan dan permasalahan utama dalam penerapan dan pengembangan pengendalian hayati adalah modal investasi permulaan yang besar yang harus dikeluarkan untuk kegiatan eksplorasi, penelitian, pengujian dan evaluasi terutama yang menyangkut berbagai aspek dasar baik untuk hama, musuh alami maupun tanaman. Aspek dasar dapat meliputi taksonomi, ekologi, biologi, siklus hidup, dinamika populasi, genetika, fisiologi, dll. Identifikasi yang tepat baik untuk jenis hama maupun musuh alaminya merupakan langkah permulaan yang sangat penting.
Kecuali diperlukan modal, fasilitas yang lengkap juga diperlukan sumber daya manusia terutama para peneliti yang berkualitas dan berpendidikan khusus dan berdedikasi tinggi sesuai dengan yang diperlukan untuk pengembangan teknologi pengendalian hayati. Meskipun ada beberapa ahli yang berpendapat bahwa untuk pengendalian hayati yang penting adalah adanya tenaga peneliti yang berpengalaman dan berdedikasi tinggi serta cukup memiliki rasa seni dan intuisi, namun bagaimanapun untuk keberhasilan pengendalian hayati dalam kerangka PHT diperlukan juga dasar pengetahuan dan teknologi yang mantap.
II. PEMBAHASAN
Adanya kekhawatiran akan pengaruh negatif tentang pemakaian agrokhemikal telah meningkatkan perhatian masyarakat kepada bioinsektisida sebagai alternative teknologi untuk menurunkan populasi hama. Salah satu alternatif pengendalian adalah pemanfaatan jamur penyebab penyakit pada serangga (bioinsectisida), yaitu jamur patogen serangga Beauveria bassiana.
Di Indonesia, hasil-hasil penelitian B. bassiana juga telah banyak dipublikasikan, terutama dari tanaman pangan untuk mengendalikan serangga hama kedelai (Riptortus linearis dan Spodoptera litura), walang sangit pada padi (Leptocoriza acuta) (Prayogo, 2006), Plutella xylostella pada sayursayuran (Hardiyanti, 2006), hama bubuk buah kopi Helopeltis antoni, dan penggerek buah kakaoHypothenemus hampei (Sudarmadji dan Prayogo, dalam Prayogo, 2006).
Saat ini produk bioinsektisida berbahan aktif B. bassiana telah tersedia secara komersial di Indonesia. Meskipun demikian, tampaknya pemanfaatannya di lapang khususnya untuk tanaman perkebunan belum optimal. Padahal,lingkungan mikro tanaman perkebunan sangat ideal bagi perkembangan epizootik cendawan-cendawan entomopatogen, termasuk B. bassiana. Keberlangsungan epizootik cendawan sangat dipengaruhi oleh suhu dan kelembaban lingkungan, dan kriteria ini dapat ditemukan pada tanaman-tanaman perkebunan yang banyak diusahakan di Indonesia. Disamping itu, pemanfaatan cendawan ini dan patogen serangga secara umum dalam pengendalian hama berpotensi memberi keuntungan ekologis jangka panjang terhadap keseimbangan hayati maupun keberlanjutan sistem pertanian.
Pengendalian penyakit tanaman dengan menggunakan agens pengendali hayati muncul karena kekhawatiran masyarakat dunia akibat penggunaan pestisida kimia sintetis. Adanya kekhawatiran tersebut membuat pengendalian hayati menjadi salah satu pilihan cara mengendalikan patogen tanaman yang harus dipertimbangkan.
Agensia hayati adalah setiap organisme yang meliputi spesies, subspesies, varietas, semua jenis serangga, nematoda, protozoa, cendawan (fungi), bakteri, virus, mikoplasma, serta organisme lainnya dalam semua tahap perkembangannya yang dapat dipergunakan untuk keperluan pengendalian hama dan penyakit atau organisme pengganggu, proses produksi, pengolahan hasil pertanian, dan berbagai keperluan lainnya (Permentan No. 411 tahun 1995).
Beauvaria bassiana merupakan cendawan entomopatogen yaitu cendawan yang dapat menimbulkan penyakit pada serangga. Beauveria bassiana secara alami terdapat di dalam tanah sebagai jamur saprofit. Pertumbuhan jamur di dalam tanah sangat dipengaruhi oleh kondisi tanah, seperti kandungan bahan organik, suhu, kelembapan, kebiasaan makan serangga, adanya pestisida sintetis, dan waktu aplikasi. Secara umum, suhu di atas 30 °C, kelembapan tanah yang berkurang dan adanya antifungal atau pestisida dapat menghambat pertumbuhannya.
Beauveria bassiana membunuh hama melalui infeksi sebagai akibat dari serangga yang kontak dengan spora jamur. Serangga dapat kontak dengan spora jamur melalui beberapa cara: semprotan jamur menempel pada tubuh serangga, serangga bergerak pada permukaan tanaman yang sudah terinfeksi jamur, atau dengan memakan jaringan tanaman yang telah diperlakukan dengan jamur. Setelah spora jamur melekat pada kulit serangga (kutikula), mereka berkecambah membentuk struktur (hifa) yang menembus tubuh serangga dan berkembang biak. Proses ini memakan waktu 3-5 hari sampai akhirnya serangga mati, bangkai yang terinfeksi dapat berfungsi sebagai sumber spora untuk penyebaran sekunder jamur. Keberhasilan penyemprotan menggunakan jamur Beauveria bassiana bergantung pada kerentanan spesies yang bersangkutan, tingkat populasi hama, dan kondisi lingkungan pada saat aplikasi, serta sumber daya manusia itu sendiri.
Mekanisme Infeksi Beauveria Bassiana
B. bassiana memproduksi toksin yang disebut beauvericin (Kučera dan Samšiňáková, 1968). Antibiotik ini dapat menyebabkan gangguan pada fungsi hemolimfa dan nukleus serangga, sehingga mengakibatkan pembengkakan yang disertai pengerasan pada serangga yang terinfeksi. Selain secara kontak, B. bassiana juga dapat menginfeksi serangga melalui inokulasi atau kontaminasi pakan. Broome et al. (1976) menyatakan bahwa 37% dari konidia B. bassiana yang dicampurkan ke dalam pakan semut api, Selenopsis richteri, berkecambah di dalam saluran pencernaan inangnya dalam waktu 72 jam, sedangkan hifanya mampu menembus dinding usus antara 60-72 jam. Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat. Serangga yang mati tidak selalu disertai gejala pertumbuhan spora. Contohnya, aphid yang terinfeksi B.bassiana hanya mengalami pembengkakan tanpa terjadi perubahan warna. Demikian pula tempayak lalat yang terinfeksi B. bassiana sering ditemukan secara berkelompok pada ujung-ujung rerumputan (Plate, 1976).
Kematian serangga biasanya disebabkan oleh kerusakan jaringan secara menyeluruh, dan atau karena toksin yang diproduksi oleh cendawan. Menurut Cheung dan Grula (1982), penyakit white muscardine yang menyerang saluran pencernaan Heliothis zea mengakibatkan gangguan nutrisi hingga kematian. Serangga yang terbunuh tubuhnya akan berwarna putih karena ditumbuhi konidia B. bassiana. Jumlah konidia yang dapat dihasilkan oleh satu serangga ditentukan oleh besar kecilnya ukuran serangga tersebut. Setiap serangga terinfeksi B. bassiana akan efektif menjadi sumber infeksi bagi serangga sehat di sekitarnya.
Seperti cendawan lain, perrtumbuhan B. bassiana juga sangat ditentukan oleh kelembapan lingkungan. Namun demikian, cendawan ini juga memiliki fase resisten yang dapat mempertahankan kemampuannya menginfeksi inang pada kondisi kering. Keberadaan epizootiknya di alam menyebabkan B. bassiana secara cepat menginfeksi populasi serangga hingga menyebabkan kematian. Selain itu, kemampuan penetrasinya yang tinggi pada tubuh serangga menyebabkan cendawan ini juga dengan mudah menginfeksi serangga hama pengisap, seperti aphid (Aphis sp.) dan kutu putih Bemisia spp. yang tidak mudah terinfeksi oleh bakteri maupun virus.
Faktor lingkungan, terutama kelembaban dan temperatur serta sedikit cahaya sangat penting perannya dalam proses infeksi dan sporulasi cendawan entomopatogen (Roberts dan Campbell, 1977; McCoy et al., 1988). Temperatur optimum untuk perkembangan, patogenisitas, dan kelulusan hidup cendawan umumnya antara 20-30°C (McCoy et al., 1988). Untuk perkecambahan konidia dan sporulasi pada permukaan tubuh serangga dibutuhkan kelembaban sangat tinggi (> 90% RH), terutama kelembaban di lingkungan mikro sekitar konidia sangat penting perannya dalam proses perkecambahan dan produksi konidia (Millstein et al., 1983; Nordin et al., 1983). Tetapi sebaliknya untuk melepaskan konidia B. bassiana dari konidifor hanya dibutuhkan kelembaban sekitar 50% (Gottwald dan Tedders, 1982).
Meskipun pengaruh cahaya terhadap infeksi cendawan belum diketahui secara jelas, tetapi intensitas sinar ultraviolet tertentu dapat merusak konidia cendawan (Callaghan, 1969). Fuxa (1987) menyatakan bahwa intensitas cahaya matahari dengan rata-rata panjang gelombang antara 290-400 nm cukup efektif menurunkan persistensi deposit konidia pada pertanaman. Sementara Ignoffo et al. (1977) mengemukakan bahwa waktu paruh (half-life) sebagian besar spora cendawan yang terekspos cahaya buatan dengan panjang gelombang mendekati panjang gelombang sinar matahari (290-400 nm) hanya sekitar 1-4 jam, tetapi kenyataannya di lapang waktu paruh dapat mencapai lebih dari 4 jam.
Umumnya cendawan entomopatogen membutuhkan lingkungan yang lembab untuk dapat menginfeksi serangga, oleh karena itu epizootiknya di alam biasanya terbentuk pada saat kondisi lingkungan lembab atau basah. Keefektifan B. bassiana menginfeksi serangga hama tergantung pada spesies atau strain cendawan, dan kepekaan stadia serangga pada tingkat kelembaban lingkungan, struktur tanah (untuk serangga dalam tanah), dan temperatur yang tepat. Selain itu, harus terjadi kontak antara spora B. bassiana yang diterbangkan angin atau terbawa air dengan serangga inang agar terjadi infeksi. Plate (1976) juga menyatakan bahwa epizootik cendawan yang terbentuk secara alami efektif mengendalikan populasi aphid, tempayak lalat yang menyerang perakaran tanaman, belalang, dan thrip, disamping juga potensial sebagai faktor mortalitas utama aphid yang menyerang kentang dan tanaman inang lainnya.
Konidia merupakan unit B. bassiana yang paling infektif dan stabil untuk aplikasi di lapang dibandingkan dengan hifa maupun blastosporanya (Soper dan Ward, 1981; Feng et al., 1994). Konidia yang diaplikasikan dapat berupa suspensi (tidak diformulasi), formulasi butiran, dan bentuk pellet, dan ketiganya memperlihatkan hasil pengendalian yang cukup nyata. Stimac et al. (1993) menyatakan bahwa aplikasi konidia B. bassiana dengan cara sprinkle dan disemprotkan pada permukaan tanah sangat efektif menyebabkan mortalitas hama sasaran. Mortalitas hama semut api, Selenopsis invicta, yang dikendalikan dengan B. bassiana tertinggi mulai 3-8 hari setelah perlakuan. Sedangkan enkapsulasi (pellet) konidia B. bassiana dengan menggunakan kalsium alginat juga efektif meningkatkan mortalitas S. invicta (White, 1995), karena enkapsulasi menyebabkan konidia lebih stabil di dalam tanah.
Beberapa senyawa metabolit sekunder diproduksi oleh B. bassiana, seperti beauvericin, bassianin, bassiacridin, bassianolide, beauverolides, tenellin, dan oosporein (Strasser et al., 2000; Vey et al., 2001; Quesada-Moraga dan Vey, 2004). Senyawa metabolit sekunder ini dapat dihasilkan oleh B. bassiana pada epizootik di alam (tanah) maupun pada epizootik buatan (di laboratorium). Meskipun demikian, hingga saat ini belum ada laporan tentang tercemarnya rantai makanan oleh senyawa metabolit sekunder, atau terakumulasi di alam sebagai limbah epizootik B. bassiana (Vey et al., 2001).
Penggunaan B. bassiana dalam pengendalian hama telah diuji secara luas di berbagai negara. Hasil uji toksikologi terhadap salah satu produk B. bassiana, Botanigard, menunjukkan bahwa produk tersebut tidak menimbulkan dampak negatif yang berhubungan dengan patogenisitas dan toksisitasnya, sehingga produk tersebut digunakan secara aman selama lebih dari 10 tahun di Amerika Serikat dan juga di beberapa negara lain (US EPA, 2006).
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna. Namun, Plate (1976) mengungkapkan bahwa tingkat kepekaan serangga bukan sasaran terhadap infeksi B. bassiana sangat ditentukan oleh virulensi dan patogenisitas cendawan, serta spesies serangga inang. Selain itu, perbedaan fisiologis dan ekologis inang juga mempengaruhi infeksi B. bassiana. Misalnya, serangga bukan sasaran yang mudah terinfeksi B. bassiana di laboratorium tidak akan serta merta terinfeksi pada kondisi lapang. Ludwig dan Oetting (2001) menegaskan bahwa beberapa serangga musuh alami yang peka terhadap infeksi B. bassiana di laboratorium ternyata mengalami infeksi sangat rendah pada uji di rumah kaca. Disamping itu, hasil uji ekotoksikologi terhadap produk Botanigard menunjukkan bahwa risiko secara ekologis yang diperlihatkan oleh serangga bukan sasaran yang diperlakukan dengan formulasi B. bassiana sangat rendah (US EPA, 2006).
Dihubungkan dengan keamanan secara hayati, cendawan entomopatogen dikelompokkan menjadi cendawan dengan kisaran inang spesifik dan yang kisaran inangnya luas (MacLeod, 1963). Cendawan yang memiliki kisaran inang spesifik umumnya menjadi parasit sejati (obligat) dan bersifat sangat virulen tarhadap inang. Sebaliknya yang kisaran inangnya luas sebagian besar merupakan patogen fakultatif, bersifat saprofit, dan cenderung kurang patogenik (Goettel et al., 1990), dan biasanya virulensinya tinggi hanya pada spesies inang dari mana cendawan tersebut pertama kali diisolasi. Contoh, B. bassiana yang diisolasi dari ulat H. armigera akan lebih patogenik pada inangnya tersebut dibanding dengan inang-inangnya yang lain. Selain itu, cendawan yang kisaran inangnya lebih luas justru menjadi lebih spesifik menginfeksi inang jika di lapang. Hal ini dapat terjadi kemungkinan karena dipengaruhi oleh interaksi antara faktor abiotik dan biotik di lapang, sehingga serangga yang mudah terinfeksi di laboratorium belum tentu mudah juga terinfeksi di lapang. Oleh karena itu, kemungkinan terinfeksinya serangga bukan sasaran oleh B. bassiana di lapang sangat kecil. Dengan demikian, aplikasi B. bassiana di lapang cenderung aman bagi musuh alami atau serangga berguna lainnya. Infeksi B. bassiana pada manusia sangat jarang terjadi. Meskipun demikian, dilaporkan ada dua kasus infeksi B. bassiana yang menyebabkan mikosis pada manusia (Henke et al., 2002; Tucker et al., 2004). Namun infeksi tersebut terjadi pada kondisi kesehatan manusia yang sangat buruk akibat penyakit leukimia akut.
Pengujian Botanigard terhadap mamalia, burung, dan ikan juga tidak menunjukkan pengaruh negatif terhadap perkembangan hewan-hewan tersebut (US EPA, 2006). Demikian pula pengujian terhadap sejumlah reptil maupun vertebrata yang membuktikan bahwa B. bassiana tidak menginfeksi keduanya (Georg et al., 1962; Fromtling et al., 1979; Gonzalez et al., 1995).
Cukup banyak tersedia bahan untuk media alami perbanyakan B. bassiana, antara lain: beras, gandum, kedelai, jagung, padi-padian, sorghum, kentang, roti, dan kacang-kacangan. Bahan mana yang akan digunakan tergantung pada beberapa faktor, termasuk kemudahan memperoleh bahan tersebut, biaya, dan strain isolat yang akan diperbanyak. Dalam perbanyakan B. bassiana dengan bahan-bahan alami, untuk menghasilkan konidia dalam jumlah maksimal diperlukan media dengan partikel yang permukaannya lebih luas. Bahan media yang cenderung menggumpal akan memiliki luas permukaan yang sempit, sehingga produksi konidia juga sedikit. Media yang ideal adalah media yang tidak hanya mempunyai partikel dengan permukaan luas, tetapi juga yang dapat mempertahankan keutuhan partikel selama proses produksi (Maheva et al., 1984; Bradley et al., 1992).
Tiga jenis bahan media alami yang telah dicoba dalam perbanyakan B. bassiana skala besar di New Zealand adalah beras, gandum, dan barley. Hasilnya, beras merupakan media paling sesuai bagi perkembangan B. bassiana dengan produktivitas konidia tertinggi mencapai 4,38 x 109 konidia/g beras (Nelson dan Glare, 1996). Penggunaan berbagai jenis sereal, selain beras, sebagai media perbanyakan B. bassiana perlu dipertimbangkan mengingat kandungan nutrisinya yang sangat bervariasi (Jenkins et al., 1998). Perbedaan kandungan nutrisi ini sangat mempengaruhi produksi konidia, terutama per kelompok produksi (batch). Oleh karena itu, pemilihan bahan media perbanyakan harus dilakukan secara tepat, terutama memilih bahan yang memiliki kemampuan produksi konidia secara konsisten dalam kelompok-kelompok produksi. Hasil penelitian lain juga membuktikan bahwa beras putih merupakan bahan media perbanyakan B. bassiana yang tepat karena produksi konidia yang tinggi (Alves dan Pereira, 1989; Mendonca, 1992; Ibrahim dan Low, 1993; Milner et al., 1993). Hal tersebut
Menunjukkan bahwa kombinasi faktor-faktor produkai sangat kompatibel, termasuk keseimbangan nutrisi dalam bahan media, biaya produksi, kemudahan memperoleh bahan, karakter fisik bahan, seperti ukuran, bentuk, dan keutuhan bahan baik sebelum maupun setelah pengkolonisasian konidia.
Dalam perbanyakan, temperatur inkubasi dan cahaya sangat menentukan produktivitas konidia. Temperatur optimal setiap cendawan bervariasi tidak saja antar spesies, tetapi juga antar isolat (Thomas dan Jenkins, 1997; Alasoadura, 1963). Temperatur optimal untuk perkecambahan konidia adalah 25-30°C, dengan temperatur minimum 10°C dan maksimum 32°C. Sedangkan pH optimal untuk pertumbuhan adalah 5,7-5,9, tapi idealnya pH 7-8 (Goral dan Lappa, 1972). Beberapa cendawan membutuhkan cahaya untuk proses sporulasi, sedangkan cendawan lainnya tidak terpengaruh oleh cahaya. Tetapi ada pula cendawan yang sporulasinya terhambat pada tingkat intensitas cahaya tertentu (Vouk dan Klas, 1931). Penelitian terdahulu membuktikan bahwa B. bassiana yang diproduksi di lingkungan tanpa cahaya (gelap) konidianya cenderung berukuran lebih besar dan lebih virulen dibanding yang diproduksi pada tempat terang (Humphreys et al., 1989; Williams, 1959). Hal ini penting sebagai bahan pertimbangan dalam memilih kemasan yang sesuai apabila biakan cendawan harus dibawa ke luar areal perbanyakan. Selain itu yang lebih penting dalam perbanyakan B. bassiana untuk skala komersial adalah kesesuaian produk dengan teknik formulasi dan aplikasinya.
Umumnya produk B. bassiana diformulasi dalam bentuk bubuk (powder) dan merupakan formulasi paling efektif memicu kontak dengan hama sasaran (Stimac et al., 1993). Formulasi B. bassiana berupa pellet hasil enkapsulasi miselium selain efektif untuk meningkatkan mortalitas hama juga untuk mengurangi kompetisi dengan mikroba lain, sehingga meningkatkan daya hidup B. bassiana (White, 1995).
Dalam jurnal diketahui kalau penambahan tepung tapioka sebanyak 1 g dan suhu penyimpanan 5OC, mempunyai potensi yang baik dalam mempertahankan viabilitas spora B. bassiana sekurang-kurangnya sampai dua bulan penyimpanan. Hal tersebut disebabkan oleh kemampuan daya simpan tepung tapioka yang lebih baik daripada tepung beras dan maizena serta didukung oleh suhu rendah yang sesuai untuk mempertahankan viabilitas spora B. bassiana. Terjadi penurunan viabilitas spora kering B. bassiana lebih cepat, seiring dengan semakin meningkatnya dosis pembawa, suhu dan lama penyimpanan. Penyimpanan pada suhu 5OC menunjukkan kemampuan mempertahankan viabilitas spora B. bassiana kering murni lebih lama daripada kondisi suhu 23C dan suhu 29C.
II. PENUTUP
Kesimpulan
Dari pembahasan di atas kita dapat menyimpulkan bahwa:
Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat
Ada beberapa faktor yang mempengaruhi mekanisme infeksi cendawan entomopatogen Beauveria bassiana, faktor yang meliputi diantaranya kelembaban, suhu, iklim, dan tingkat kekebalan tubuh dari inang
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna.
DAFTAR PUSTAKA
Anonim, 2012. Perbanyakan Cendawan Entomopatogen. http: www. Pengendalian%agents-hayati.com.html.diakses pada tanggal 21 maret 2012
Anonim, 2012. Prospek Beauveria bassiana sebagai bioinsectisida. http//:www. Miss Biology Prospek Beauveria bassiana sebagai bioinsectisida.htm. diakses pada tanggal 21 maret 2012
Anonim, 2012. Beauveria bassiana . Wikipedia bahasa Indonesia, ensiklopedia bebas.html. diakses pada tanggal 21 maret 2012
Balai Penelitian Tanaman Hias. 2006. Beauveria bassiana pengendali hama tanaman. Warta Penelitian dan Pengembangan Pertanian Indonesia 28(1): 11-12.
EOL. 2000. Beauveria bassiana Bals. -Criv. Vuill. 1912.[terhubung berkala] http://www.eol.org/pages/160292 . diakses pada tanggal 21 maret 2012
McCoy C, Quintela ED, Faria M de. 1990. Environmental persistence of entomopathogenic fungi. [terhubung berkala]. http://www.lsuagcenter.com/s265/mccoy.htm. diakses pada tanggal 21 maret 2012
I. PENDAHULUAN
Agens Pengendali Hayati adalah musuh alami berupa Parasitoid, Predator, Entomopatogen dan Agens Antagonis yang telah diketahui mempunyai potensi yang besar sebagai salah satu sarana pengendalian OPT.
Entomopatogen atau penyakit pada serangga hama yang mencakup semua mikroba yaitu Cendawan, bakteri, virus, dan nematoda yang memiliki kemampuan untuk menjadi parasit (pathogen) pada serangga sehingga dapat mematikan serangga yang diinfeksinya. Cendawan adalah mikroba yang mempunyai bentuk tubuh menyerupai benang yang disebut hifa, nampak seperti tumbuhan namun tidak memiliki klorofil.
Mikroba dari jenis jamur adalah Beauveria bassiana, Metarrhizium anisopliae Metarrhizium ftavoviridae Verticillium lecanii dll. dapat mengendalikan Uret, Kepinding tanah, Belalang, Lalat buah dan serangga lainnya.
Kesulitan dan permasalahan utama dalam penerapan dan pengembangan pengendalian hayati adalah modal investasi permulaan yang besar yang harus dikeluarkan untuk kegiatan eksplorasi, penelitian, pengujian dan evaluasi terutama yang menyangkut berbagai aspek dasar baik untuk hama, musuh alami maupun tanaman. Aspek dasar dapat meliputi taksonomi, ekologi, biologi, siklus hidup, dinamika populasi, genetika, fisiologi, dll. Identifikasi yang tepat baik untuk jenis hama maupun musuh alaminya merupakan langkah permulaan yang sangat penting.
Kecuali diperlukan modal, fasilitas yang lengkap juga diperlukan sumber daya manusia terutama para peneliti yang berkualitas dan berpendidikan khusus dan berdedikasi tinggi sesuai dengan yang diperlukan untuk pengembangan teknologi pengendalian hayati. Meskipun ada beberapa ahli yang berpendapat bahwa untuk pengendalian hayati yang penting adalah adanya tenaga peneliti yang berpengalaman dan berdedikasi tinggi serta cukup memiliki rasa seni dan intuisi, namun bagaimanapun untuk keberhasilan pengendalian hayati dalam kerangka PHT diperlukan juga dasar pengetahuan dan teknologi yang mantap.
II. PEMBAHASAN
Adanya kekhawatiran akan pengaruh negatif tentang pemakaian agrokhemikal telah meningkatkan perhatian masyarakat kepada bioinsektisida sebagai alternative teknologi untuk menurunkan populasi hama. Salah satu alternatif pengendalian adalah pemanfaatan jamur penyebab penyakit pada serangga (bioinsectisida), yaitu jamur patogen serangga Beauveria bassiana.
Di Indonesia, hasil-hasil penelitian B. bassiana juga telah banyak dipublikasikan, terutama dari tanaman pangan untuk mengendalikan serangga hama kedelai (Riptortus linearis dan Spodoptera litura), walang sangit pada padi (Leptocoriza acuta) (Prayogo, 2006), Plutella xylostella pada sayursayuran (Hardiyanti, 2006), hama bubuk buah kopi Helopeltis antoni, dan penggerek buah kakaoHypothenemus hampei (Sudarmadji dan Prayogo, dalam Prayogo, 2006).
Saat ini produk bioinsektisida berbahan aktif B. bassiana telah tersedia secara komersial di Indonesia. Meskipun demikian, tampaknya pemanfaatannya di lapang khususnya untuk tanaman perkebunan belum optimal. Padahal,lingkungan mikro tanaman perkebunan sangat ideal bagi perkembangan epizootik cendawan-cendawan entomopatogen, termasuk B. bassiana. Keberlangsungan epizootik cendawan sangat dipengaruhi oleh suhu dan kelembaban lingkungan, dan kriteria ini dapat ditemukan pada tanaman-tanaman perkebunan yang banyak diusahakan di Indonesia. Disamping itu, pemanfaatan cendawan ini dan patogen serangga secara umum dalam pengendalian hama berpotensi memberi keuntungan ekologis jangka panjang terhadap keseimbangan hayati maupun keberlanjutan sistem pertanian.
Pengendalian penyakit tanaman dengan menggunakan agens pengendali hayati muncul karena kekhawatiran masyarakat dunia akibat penggunaan pestisida kimia sintetis. Adanya kekhawatiran tersebut membuat pengendalian hayati menjadi salah satu pilihan cara mengendalikan patogen tanaman yang harus dipertimbangkan.
Agensia hayati adalah setiap organisme yang meliputi spesies, subspesies, varietas, semua jenis serangga, nematoda, protozoa, cendawan (fungi), bakteri, virus, mikoplasma, serta organisme lainnya dalam semua tahap perkembangannya yang dapat dipergunakan untuk keperluan pengendalian hama dan penyakit atau organisme pengganggu, proses produksi, pengolahan hasil pertanian, dan berbagai keperluan lainnya (Permentan No. 411 tahun 1995).
Beauvaria bassiana merupakan cendawan entomopatogen yaitu cendawan yang dapat menimbulkan penyakit pada serangga. Beauveria bassiana secara alami terdapat di dalam tanah sebagai jamur saprofit. Pertumbuhan jamur di dalam tanah sangat dipengaruhi oleh kondisi tanah, seperti kandungan bahan organik, suhu, kelembapan, kebiasaan makan serangga, adanya pestisida sintetis, dan waktu aplikasi. Secara umum, suhu di atas 30 °C, kelembapan tanah yang berkurang dan adanya antifungal atau pestisida dapat menghambat pertumbuhannya.
Beauveria bassiana membunuh hama melalui infeksi sebagai akibat dari serangga yang kontak dengan spora jamur. Serangga dapat kontak dengan spora jamur melalui beberapa cara: semprotan jamur menempel pada tubuh serangga, serangga bergerak pada permukaan tanaman yang sudah terinfeksi jamur, atau dengan memakan jaringan tanaman yang telah diperlakukan dengan jamur. Setelah spora jamur melekat pada kulit serangga (kutikula), mereka berkecambah membentuk struktur (hifa) yang menembus tubuh serangga dan berkembang biak. Proses ini memakan waktu 3-5 hari sampai akhirnya serangga mati, bangkai yang terinfeksi dapat berfungsi sebagai sumber spora untuk penyebaran sekunder jamur. Keberhasilan penyemprotan menggunakan jamur Beauveria bassiana bergantung pada kerentanan spesies yang bersangkutan, tingkat populasi hama, dan kondisi lingkungan pada saat aplikasi, serta sumber daya manusia itu sendiri.
Mekanisme Infeksi Beauveria Bassiana
B. bassiana memproduksi toksin yang disebut beauvericin (Kučera dan Samšiňáková, 1968). Antibiotik ini dapat menyebabkan gangguan pada fungsi hemolimfa dan nukleus serangga, sehingga mengakibatkan pembengkakan yang disertai pengerasan pada serangga yang terinfeksi. Selain secara kontak, B. bassiana juga dapat menginfeksi serangga melalui inokulasi atau kontaminasi pakan. Broome et al. (1976) menyatakan bahwa 37% dari konidia B. bassiana yang dicampurkan ke dalam pakan semut api, Selenopsis richteri, berkecambah di dalam saluran pencernaan inangnya dalam waktu 72 jam, sedangkan hifanya mampu menembus dinding usus antara 60-72 jam. Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat. Serangga yang mati tidak selalu disertai gejala pertumbuhan spora. Contohnya, aphid yang terinfeksi B.bassiana hanya mengalami pembengkakan tanpa terjadi perubahan warna. Demikian pula tempayak lalat yang terinfeksi B. bassiana sering ditemukan secara berkelompok pada ujung-ujung rerumputan (Plate, 1976).
Kematian serangga biasanya disebabkan oleh kerusakan jaringan secara menyeluruh, dan atau karena toksin yang diproduksi oleh cendawan. Menurut Cheung dan Grula (1982), penyakit white muscardine yang menyerang saluran pencernaan Heliothis zea mengakibatkan gangguan nutrisi hingga kematian. Serangga yang terbunuh tubuhnya akan berwarna putih karena ditumbuhi konidia B. bassiana. Jumlah konidia yang dapat dihasilkan oleh satu serangga ditentukan oleh besar kecilnya ukuran serangga tersebut. Setiap serangga terinfeksi B. bassiana akan efektif menjadi sumber infeksi bagi serangga sehat di sekitarnya.
Seperti cendawan lain, perrtumbuhan B. bassiana juga sangat ditentukan oleh kelembapan lingkungan. Namun demikian, cendawan ini juga memiliki fase resisten yang dapat mempertahankan kemampuannya menginfeksi inang pada kondisi kering. Keberadaan epizootiknya di alam menyebabkan B. bassiana secara cepat menginfeksi populasi serangga hingga menyebabkan kematian. Selain itu, kemampuan penetrasinya yang tinggi pada tubuh serangga menyebabkan cendawan ini juga dengan mudah menginfeksi serangga hama pengisap, seperti aphid (Aphis sp.) dan kutu putih Bemisia spp. yang tidak mudah terinfeksi oleh bakteri maupun virus.
Faktor lingkungan, terutama kelembaban dan temperatur serta sedikit cahaya sangat penting perannya dalam proses infeksi dan sporulasi cendawan entomopatogen (Roberts dan Campbell, 1977; McCoy et al., 1988). Temperatur optimum untuk perkembangan, patogenisitas, dan kelulusan hidup cendawan umumnya antara 20-30°C (McCoy et al., 1988). Untuk perkecambahan konidia dan sporulasi pada permukaan tubuh serangga dibutuhkan kelembaban sangat tinggi (> 90% RH), terutama kelembaban di lingkungan mikro sekitar konidia sangat penting perannya dalam proses perkecambahan dan produksi konidia (Millstein et al., 1983; Nordin et al., 1983). Tetapi sebaliknya untuk melepaskan konidia B. bassiana dari konidifor hanya dibutuhkan kelembaban sekitar 50% (Gottwald dan Tedders, 1982).
Meskipun pengaruh cahaya terhadap infeksi cendawan belum diketahui secara jelas, tetapi intensitas sinar ultraviolet tertentu dapat merusak konidia cendawan (Callaghan, 1969). Fuxa (1987) menyatakan bahwa intensitas cahaya matahari dengan rata-rata panjang gelombang antara 290-400 nm cukup efektif menurunkan persistensi deposit konidia pada pertanaman. Sementara Ignoffo et al. (1977) mengemukakan bahwa waktu paruh (half-life) sebagian besar spora cendawan yang terekspos cahaya buatan dengan panjang gelombang mendekati panjang gelombang sinar matahari (290-400 nm) hanya sekitar 1-4 jam, tetapi kenyataannya di lapang waktu paruh dapat mencapai lebih dari 4 jam.
Umumnya cendawan entomopatogen membutuhkan lingkungan yang lembab untuk dapat menginfeksi serangga, oleh karena itu epizootiknya di alam biasanya terbentuk pada saat kondisi lingkungan lembab atau basah. Keefektifan B. bassiana menginfeksi serangga hama tergantung pada spesies atau strain cendawan, dan kepekaan stadia serangga pada tingkat kelembaban lingkungan, struktur tanah (untuk serangga dalam tanah), dan temperatur yang tepat. Selain itu, harus terjadi kontak antara spora B. bassiana yang diterbangkan angin atau terbawa air dengan serangga inang agar terjadi infeksi. Plate (1976) juga menyatakan bahwa epizootik cendawan yang terbentuk secara alami efektif mengendalikan populasi aphid, tempayak lalat yang menyerang perakaran tanaman, belalang, dan thrip, disamping juga potensial sebagai faktor mortalitas utama aphid yang menyerang kentang dan tanaman inang lainnya.
Konidia merupakan unit B. bassiana yang paling infektif dan stabil untuk aplikasi di lapang dibandingkan dengan hifa maupun blastosporanya (Soper dan Ward, 1981; Feng et al., 1994). Konidia yang diaplikasikan dapat berupa suspensi (tidak diformulasi), formulasi butiran, dan bentuk pellet, dan ketiganya memperlihatkan hasil pengendalian yang cukup nyata. Stimac et al. (1993) menyatakan bahwa aplikasi konidia B. bassiana dengan cara sprinkle dan disemprotkan pada permukaan tanah sangat efektif menyebabkan mortalitas hama sasaran. Mortalitas hama semut api, Selenopsis invicta, yang dikendalikan dengan B. bassiana tertinggi mulai 3-8 hari setelah perlakuan. Sedangkan enkapsulasi (pellet) konidia B. bassiana dengan menggunakan kalsium alginat juga efektif meningkatkan mortalitas S. invicta (White, 1995), karena enkapsulasi menyebabkan konidia lebih stabil di dalam tanah.
Beberapa senyawa metabolit sekunder diproduksi oleh B. bassiana, seperti beauvericin, bassianin, bassiacridin, bassianolide, beauverolides, tenellin, dan oosporein (Strasser et al., 2000; Vey et al., 2001; Quesada-Moraga dan Vey, 2004). Senyawa metabolit sekunder ini dapat dihasilkan oleh B. bassiana pada epizootik di alam (tanah) maupun pada epizootik buatan (di laboratorium). Meskipun demikian, hingga saat ini belum ada laporan tentang tercemarnya rantai makanan oleh senyawa metabolit sekunder, atau terakumulasi di alam sebagai limbah epizootik B. bassiana (Vey et al., 2001).
Penggunaan B. bassiana dalam pengendalian hama telah diuji secara luas di berbagai negara. Hasil uji toksikologi terhadap salah satu produk B. bassiana, Botanigard, menunjukkan bahwa produk tersebut tidak menimbulkan dampak negatif yang berhubungan dengan patogenisitas dan toksisitasnya, sehingga produk tersebut digunakan secara aman selama lebih dari 10 tahun di Amerika Serikat dan juga di beberapa negara lain (US EPA, 2006).
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna. Namun, Plate (1976) mengungkapkan bahwa tingkat kepekaan serangga bukan sasaran terhadap infeksi B. bassiana sangat ditentukan oleh virulensi dan patogenisitas cendawan, serta spesies serangga inang. Selain itu, perbedaan fisiologis dan ekologis inang juga mempengaruhi infeksi B. bassiana. Misalnya, serangga bukan sasaran yang mudah terinfeksi B. bassiana di laboratorium tidak akan serta merta terinfeksi pada kondisi lapang. Ludwig dan Oetting (2001) menegaskan bahwa beberapa serangga musuh alami yang peka terhadap infeksi B. bassiana di laboratorium ternyata mengalami infeksi sangat rendah pada uji di rumah kaca. Disamping itu, hasil uji ekotoksikologi terhadap produk Botanigard menunjukkan bahwa risiko secara ekologis yang diperlihatkan oleh serangga bukan sasaran yang diperlakukan dengan formulasi B. bassiana sangat rendah (US EPA, 2006).
Dihubungkan dengan keamanan secara hayati, cendawan entomopatogen dikelompokkan menjadi cendawan dengan kisaran inang spesifik dan yang kisaran inangnya luas (MacLeod, 1963). Cendawan yang memiliki kisaran inang spesifik umumnya menjadi parasit sejati (obligat) dan bersifat sangat virulen tarhadap inang. Sebaliknya yang kisaran inangnya luas sebagian besar merupakan patogen fakultatif, bersifat saprofit, dan cenderung kurang patogenik (Goettel et al., 1990), dan biasanya virulensinya tinggi hanya pada spesies inang dari mana cendawan tersebut pertama kali diisolasi. Contoh, B. bassiana yang diisolasi dari ulat H. armigera akan lebih patogenik pada inangnya tersebut dibanding dengan inang-inangnya yang lain. Selain itu, cendawan yang kisaran inangnya lebih luas justru menjadi lebih spesifik menginfeksi inang jika di lapang. Hal ini dapat terjadi kemungkinan karena dipengaruhi oleh interaksi antara faktor abiotik dan biotik di lapang, sehingga serangga yang mudah terinfeksi di laboratorium belum tentu mudah juga terinfeksi di lapang. Oleh karena itu, kemungkinan terinfeksinya serangga bukan sasaran oleh B. bassiana di lapang sangat kecil. Dengan demikian, aplikasi B. bassiana di lapang cenderung aman bagi musuh alami atau serangga berguna lainnya. Infeksi B. bassiana pada manusia sangat jarang terjadi. Meskipun demikian, dilaporkan ada dua kasus infeksi B. bassiana yang menyebabkan mikosis pada manusia (Henke et al., 2002; Tucker et al., 2004). Namun infeksi tersebut terjadi pada kondisi kesehatan manusia yang sangat buruk akibat penyakit leukimia akut.
Pengujian Botanigard terhadap mamalia, burung, dan ikan juga tidak menunjukkan pengaruh negatif terhadap perkembangan hewan-hewan tersebut (US EPA, 2006). Demikian pula pengujian terhadap sejumlah reptil maupun vertebrata yang membuktikan bahwa B. bassiana tidak menginfeksi keduanya (Georg et al., 1962; Fromtling et al., 1979; Gonzalez et al., 1995).
Cukup banyak tersedia bahan untuk media alami perbanyakan B. bassiana, antara lain: beras, gandum, kedelai, jagung, padi-padian, sorghum, kentang, roti, dan kacang-kacangan. Bahan mana yang akan digunakan tergantung pada beberapa faktor, termasuk kemudahan memperoleh bahan tersebut, biaya, dan strain isolat yang akan diperbanyak. Dalam perbanyakan B. bassiana dengan bahan-bahan alami, untuk menghasilkan konidia dalam jumlah maksimal diperlukan media dengan partikel yang permukaannya lebih luas. Bahan media yang cenderung menggumpal akan memiliki luas permukaan yang sempit, sehingga produksi konidia juga sedikit. Media yang ideal adalah media yang tidak hanya mempunyai partikel dengan permukaan luas, tetapi juga yang dapat mempertahankan keutuhan partikel selama proses produksi (Maheva et al., 1984; Bradley et al., 1992).
Tiga jenis bahan media alami yang telah dicoba dalam perbanyakan B. bassiana skala besar di New Zealand adalah beras, gandum, dan barley. Hasilnya, beras merupakan media paling sesuai bagi perkembangan B. bassiana dengan produktivitas konidia tertinggi mencapai 4,38 x 109 konidia/g beras (Nelson dan Glare, 1996). Penggunaan berbagai jenis sereal, selain beras, sebagai media perbanyakan B. bassiana perlu dipertimbangkan mengingat kandungan nutrisinya yang sangat bervariasi (Jenkins et al., 1998). Perbedaan kandungan nutrisi ini sangat mempengaruhi produksi konidia, terutama per kelompok produksi (batch). Oleh karena itu, pemilihan bahan media perbanyakan harus dilakukan secara tepat, terutama memilih bahan yang memiliki kemampuan produksi konidia secara konsisten dalam kelompok-kelompok produksi. Hasil penelitian lain juga membuktikan bahwa beras putih merupakan bahan media perbanyakan B. bassiana yang tepat karena produksi konidia yang tinggi (Alves dan Pereira, 1989; Mendonca, 1992; Ibrahim dan Low, 1993; Milner et al., 1993). Hal tersebut
Menunjukkan bahwa kombinasi faktor-faktor produkai sangat kompatibel, termasuk keseimbangan nutrisi dalam bahan media, biaya produksi, kemudahan memperoleh bahan, karakter fisik bahan, seperti ukuran, bentuk, dan keutuhan bahan baik sebelum maupun setelah pengkolonisasian konidia.
Dalam perbanyakan, temperatur inkubasi dan cahaya sangat menentukan produktivitas konidia. Temperatur optimal setiap cendawan bervariasi tidak saja antar spesies, tetapi juga antar isolat (Thomas dan Jenkins, 1997; Alasoadura, 1963). Temperatur optimal untuk perkecambahan konidia adalah 25-30°C, dengan temperatur minimum 10°C dan maksimum 32°C. Sedangkan pH optimal untuk pertumbuhan adalah 5,7-5,9, tapi idealnya pH 7-8 (Goral dan Lappa, 1972). Beberapa cendawan membutuhkan cahaya untuk proses sporulasi, sedangkan cendawan lainnya tidak terpengaruh oleh cahaya. Tetapi ada pula cendawan yang sporulasinya terhambat pada tingkat intensitas cahaya tertentu (Vouk dan Klas, 1931). Penelitian terdahulu membuktikan bahwa B. bassiana yang diproduksi di lingkungan tanpa cahaya (gelap) konidianya cenderung berukuran lebih besar dan lebih virulen dibanding yang diproduksi pada tempat terang (Humphreys et al., 1989; Williams, 1959). Hal ini penting sebagai bahan pertimbangan dalam memilih kemasan yang sesuai apabila biakan cendawan harus dibawa ke luar areal perbanyakan. Selain itu yang lebih penting dalam perbanyakan B. bassiana untuk skala komersial adalah kesesuaian produk dengan teknik formulasi dan aplikasinya.
Umumnya produk B. bassiana diformulasi dalam bentuk bubuk (powder) dan merupakan formulasi paling efektif memicu kontak dengan hama sasaran (Stimac et al., 1993). Formulasi B. bassiana berupa pellet hasil enkapsulasi miselium selain efektif untuk meningkatkan mortalitas hama juga untuk mengurangi kompetisi dengan mikroba lain, sehingga meningkatkan daya hidup B. bassiana (White, 1995).
Dalam jurnal diketahui kalau penambahan tepung tapioka sebanyak 1 g dan suhu penyimpanan 5OC, mempunyai potensi yang baik dalam mempertahankan viabilitas spora B. bassiana sekurang-kurangnya sampai dua bulan penyimpanan. Hal tersebut disebabkan oleh kemampuan daya simpan tepung tapioka yang lebih baik daripada tepung beras dan maizena serta didukung oleh suhu rendah yang sesuai untuk mempertahankan viabilitas spora B. bassiana. Terjadi penurunan viabilitas spora kering B. bassiana lebih cepat, seiring dengan semakin meningkatnya dosis pembawa, suhu dan lama penyimpanan. Penyimpanan pada suhu 5OC menunjukkan kemampuan mempertahankan viabilitas spora B. bassiana kering murni lebih lama daripada kondisi suhu 23C dan suhu 29C.
II. PENUTUP
Kesimpulan
Dari pembahasan di atas kita dapat menyimpulkan bahwa:
Di dalam tubuh inangnya cendawan ini dengan cepat memperbanyak diri hingga seluruh jaringan serangga terinfeksi. Serangga yang telah terinfeksi B. bassiana biasanya akan berhenti makan, sehingga menjadi lemah, dan kematiannya bisa lebih cepat
Ada beberapa faktor yang mempengaruhi mekanisme infeksi cendawan entomopatogen Beauveria bassiana, faktor yang meliputi diantaranya kelembaban, suhu, iklim, dan tingkat kekebalan tubuh dari inang
Cendawan B. bassiana memiliki kisaran inang sangat luas, sehingga kurang selektif terhadap inang sasaran. Hal ini memungkinkan B. bassiana dapat menginfeksi serangga bukan sasaran atau serangga berguna.
DAFTAR PUSTAKA
Anonim, 2012. Perbanyakan Cendawan Entomopatogen. http: www. Pengendalian%agents-hayati.com.html.diakses pada tanggal 21 maret 2012
Anonim, 2012. Prospek Beauveria bassiana sebagai bioinsectisida. http//:www. Miss Biology Prospek Beauveria bassiana sebagai bioinsectisida.htm. diakses pada tanggal 21 maret 2012
Anonim, 2012. Beauveria bassiana . Wikipedia bahasa Indonesia, ensiklopedia bebas.html. diakses pada tanggal 21 maret 2012
Balai Penelitian Tanaman Hias. 2006. Beauveria bassiana pengendali hama tanaman. Warta Penelitian dan Pengembangan Pertanian Indonesia 28(1): 11-12.
EOL. 2000. Beauveria bassiana Bals. -Criv. Vuill. 1912.[terhubung berkala] http://www.eol.org/pages/160292 . diakses pada tanggal 21 maret 2012
McCoy C, Quintela ED, Faria M de. 1990. Environmental persistence of entomopathogenic fungi. [terhubung berkala]. http://www.lsuagcenter.com/s265/mccoy.htm. diakses pada tanggal 21 maret 2012
Minggu, 30 Oktober 2011
Sistem Pertanian Terpadu Ubi Kayu
PENDAHULUAN
Tanaman ubi kayu (Manihotutikisima) memiliki berbagai varietas atau klon yang dapat langsung dikonsumsi sebagai makanan atau menjadi bahan baku bagi industri tapioka dan gapiek (manihok) ataupun tepung gapiek, yang selanjutnya dipergunakan untuk berbagai macam industri seperti makanan, makanan ternak, kertas, kayu lapis dan tainnya. Berdasarkan potensi fisik seperti kesesuaian lahan, iklim, sumberdaya manusia,dan tingkat adaptasi teknologi, tanaman ubi kayu banyak didapatkan dan bisa dibudidayakan di banyak tempat atau lokasi di Indonesia sehingga memungkinkan untuk diusahakan oleh para petani secara luas.
Hasil olahan ubi kayu berupa tapioka dan gapiek (manihok) dalam bentuk chips, pellet ataupun lainnya, telah lama menjadi komoditi ekspor yang sangat penting dalam menyumbang pendapatan devisa, karenanya merupakan aset yang sangat berharga dan perlu dijaga kelestariannya sehingga dapat dimanfaatkan untuk pengembangan ekspor pada masa-masa selanjutnya. Pengalaman petani menunjukkan bahwa penanaman ubi kayu sering tidak membuahkan hasil yang cukup baik untuk keluarga, karena keadaan tata niaga kayu yang banyak dipengaruhi oleh fluktuasi harga sehingga merugikan petani. Pada saat menjelang tanam, harga ubi kayu biasanya terlihat sangat menarik sehingga banyak petani berusaha menanamnya. Akan tetapi pada saat panen harga kayu kemudian jatuh sehingga banyak merugikan petani (berdasarkan data yang ada kapasitas pabrik dan potensi ekspor masih lebih besar dari jumlah produksi).
Kondisi ini telah mendorong banyak petani untuk mengalihkan perhatian dan berusaha menanam komoditi seperti kelapa sawit. Di sisi lain, apabila hal ini terus dibiarkan, akan bisa berakibat turunnya produksi yang dapat menekan pasokan ubi kayu baik untuk keperluan ekspor ataupun konsumsi sawit dalam negeri. Lemahnya posisi petani ubi kayu dalam menghadapi pengaruh fluktuasi harga, terutama disebabkan karena ubi kayu memiliki daya simpan yang rendah,dan produktifitasnya juga rendah akibat modal usaha yang sangat terbatas, disamping kebutuhan keluarga yang sudah sangat mendesak. Pendapatan petani ubi kayu akan makin rendah lagi Karen apada saat dijual ke pabrik mendapatkan mutu ubi kayunya rendah dan rafaksi yang ditentukan secara sepihak oleh pabrik.
Berdasarkan keadaan yang ada, maka salah satu alternatif untuk meng antisipasi masalah tersebut di atas, adalah dengan upaya mengadakan kerjasama antara para petani produsen ubi kayu di satu pihak dengan pengguna atau pemakai ubi kayu (baik dengan kapasitas pabrik yang cukup besar, maupun dengan kapasitas pabrik yang sedang) dipihak lain, sehingga dapat diciptakan bentuk kerjasama kemitraan dengan tata niaga yang saling menguntungkan bagi kedua belah pihak. Untuk mendukung kelancaran kemitraan ini, para petani memerlukan dukungan permodalan untuk keperluan penanaman ubi kayu, di mana bank pelaksana dapat berperan dengan memberikan kredit dengan skim kredit yang sesuai.Secara teknis upaya mengembangkan budidaya ubi kayu harus diikuti dengan adanya upaya untuk mengubah atau memperbaiki teknik budidaya yang selama ini masih konvensional menjadi teknik budidaya yang intensif. Hal ini dapat dicapai dengan lebih memantapkan penataan teknologi produksi mulai dari persiapan lahan, pengolahan lahan, penggunaan varietas unggul, pemeliharaan tanaman, Panen dan pasca panen serta distribusi pemasaran hasil.
TINJAUAN PUSTAKA
Identifikasi Potensi Sumber Daya Alam Tanaman Ubi Kayu
Ubi kayu termasuk tanaman tropis, tetapi dapat pula beradaptasi dan tumbuh dengan baik di daerah subtropis. Secara umum tanaman ini tidak menuntut iklim yang spesifik untuk pertumbuhannya. Namun demikian ubi kayu akan tumbuh dengan baik pada iklim dan tanah. iklim yang cocok untuk tanaman ubi kayu antara lain :
Curah hujan : 750 - 1.000 mm/thn
Tinggi tempat : 0 - 1.500 m dpl
Suhu : 25 ͦ - 28 ͦ C
Fungsi singkong (ubi kayu) sudah mulai bergeser, dari penyediaan bahan pangan, berpotensi menjadi bahan baku untuk pengembangan bio-ethanol. Kebutuhan bio-ethanol sampai dengan 2010 tergolong cukup tinggi, yaitu mencapai 1,8 juta kilo liter.
Pati ataun Amilum
Pati atau amilum adalah karbohidrat kompleks yang tidak larut dalam air, berwujud bubuk putih, tawar dan tidak berbau. Pati merupakan bahan utama yang dihasilkan oleh tumbuhan untuk menyimpan kelebihan glukosa (sebagai produk fotosintesis) dalam jangka panjang. Hewan dan manusia juga menjadikan pati sebagai sumber energi yang penting.
Pati tersusun dari dua macam karbohidrat, amilosa dan amilopektin, dalam komposisi yang berbeda-beda. Amilosa memberikan sifat keras (pera) sedangkan amilopektin menyebabkan sifat lengket. Amilosa memberikan warna ungu pekat pada tes iodin sedangkan amilopektin tidak bereaksi.
“Pati” (bahasa Inggris starch) adalah penyusun (utama) tepung. Tepung bisa jadi tidak murni hanya mengandung pati, karena tercampur dengan protein, pengawet, dan sebagainya. Tepung beras mengandung pati beras, protein, vitamin, dan lain-lain bahan yang terkandung pada butir beras. Orang bisa juga mendapatkan tepung yang merupakan campuran dua atau lebih pati. Kata ‘tepung lebih berkaitan dengan komoditas ekonomis. Pati digunakan sebagai bahan yang digunakan untuk memekatkan makanan cair seperti sup dan sebagainya. Dalam industri, pati dipakai sebagai komponen perekat, campuran kertas dan tekstil, dan pada industri kosmetik.
Bioetanol
Bioetanol merupakan bahan bakar dari minyak nabati yang memiliki sifat menyerupai minyak premium. Untuk pengganti premium, terdapat alternatif gasohol yang merupakan campuran antara bensin dan bioetanol. Adapun manfaat pemakaian gasohol di Indonesia yaitu memperbesar basis sumber daya bahan bakar cair, mengurangi impor BBM, menguatkan security of supply bahan bakar, meningkatkan kesempatan kerja, berpotensi mengurangi ketimpangan pendapatan antar individu dan antar daerah, meningkatkan kemampuan nasional dalam teknologi pertanian dan industri, mengurangi kecenderungan pemanasan global dan pencemaran udara (bahan bakar ramah lingkungan) dan berpotensi mendorong ekspor komoditi baru. Bioetanol tersebut bersumber dari karbohidrat yang potensial sebagai bahan baku seperti jagung, ubi kayu, ubi jalar, sagu dan tebu. Adapun konversi biomasa tanaman tersebut menjadi bioethanol adalah seperti pada tabel dibawah ini.
Tabel Konversi biomasa menjadi bioethanol
Biomassa Jumlah biomassa (kg) Kandungan gula (kg) Jumlah hasil bioetanol (liter) Biomassa : Bioetanol
Ubi Kayu 1.000 250-300 166,6 6,5 : 1
Ubi Jalar 1.000 150-200 125 8 : 1
Jagung 1.000 600-700 400 2,5 : 1
Sagu 1.000 120-160 90 12:1
Tetes 1.000 500 250 4:1
Sumber Daya Tanaman
Potensi pengembangan ubi kayu sebagai bahan bakar industri bioetanol
berdasarkan aspek biofisik di antaranya :
Produktivitas
Rataan produktivitas ubi kayu secara nasional selama dasawarsa terakhir masih rendah, yaitu sekitar 40% dari potensi genetis dengan pengelolaan optimal (BPS 2005; Howeler 2001). Faktor utama yang menyebabkan potensi hasil tinggi adalah kemampuan menghasilkan fotosintat tinggi dan periode pertumbuhannya lama. Tanaman dapat memanfaatkan sinar matahari secara maksimal melalui pengaturan deklinasi dan inklinasi tangkai daun dan helaian daun (Hozyo et al. 1984), di mana helaian daun dapat bergerak mengikuti posisi matahari, sejak pagi hingga sore, ataupun mengikuti pergerakan matahari dari Lintang Selatan ke Lintang Utara dan sebaliknya. Potensi hasil tiap varietas bersifat spesifik lokasi (Suhartina 2005). Oleh karena itu, varietas unggul seperti Adira-4, Malang-6, UJ-3, dan UJ-5 wilayah berkembangnya berbeda. Peningkatan produksi dapat direalisasikan melalui penggunaan varietas unggul dan pengelolaan optimal. Dengan demikian, pengembangan industri pada berbagai tipe agroekologi, ketersediaan bakunya berpotensi dapat dijamin.
Fleksibilitas umur panen
Hasil yang diharapkan dalam pengembangan ubi kayu sebagai bahan baku industri bioetanol adalah ubi berkadar pati tinggi untuk meningkatkan efisiensi industri. Oleh karena itu, kriteria yang harus dipenuhi dalam penentuan umur panen adalah kadar pati optimal. Kadar pati optimal dicapai pada umur yang bervariasi dan secara umum dapat dibedakan menjadi tiga kelompok, yaitu umur: (a) tujuh bulan (genjah), (b) sembilan bulan (sedang), dan (c) 10 bulan (dalam). Kualitas pati tidak berubah walaupun panennya ditunda, sedangkan bobot ubi meningkat sejalan dengan umur. Dengan demikian umur panen dapat disesuaikan dengan kebutuhan industri tanpa penurunan kualitas pati. Kondisi umur panen yang fleksibel tersebut memberikan gambaran adanya potensi untuk menjamin ketersediaan bahan baku industri, sehingga potensial dikembangkan.
Fleksibel dalam usaha tani
Ubi sebagai hasil usaha tani mempunyai peran multi guna, seperti sebagai bahan pangan, industri, pakan dan komoditas ekspor. Oleh karena itu, usaha tani ubi kayu dapat dilakukan secara subsisten, semi komersial dan komersial. Usaha tani subsisten dilakukan petani dengan tujuan utama pemenuhan kebutuhan pangan, sedangkan usaha tani semi komersial dan komersial dengan tujuan utama mendapatkan keuntungan dan pemenuhan permintaan pasar. Ubi kayu pertumbannya lambat selama tiga bulan pertama, sehingga usaha tani sistem tumpangsari dengan padi dan palawija lainnya juga potensial dikembangkan. Ubi kayu dapat dipanen mulai umur tujuh sampai 11 bulan, sehingga dapat dikategorikan sebagai tanaman semusim, dan dapat dipanen pada umur 12 bulan atau lebih, sehingga dapat dikategorikan sebagai tanaman tahunan/tanaman perkebunan. Sifat-sifat tersebut dapat digunakan sebagai indikator bahwa usaha tani ubi kayu fleksibel, dan dalam pengembangannya berpotensi menjamin ketersediaan bahan baku industri bioetanol, pangan, pakan, dan ekspor.
Efisiensi energi tinggi
Sumber energi untuk bioetanol yang berbahan baku ubi kayu adalah pati. Kadar pati varietas unggul yang tersedia berkisar 25–31%. Kadar pati tidak menurun kualitasnya walaupun panennya dilakukan setelah umur optimal, bahkan hasilnya meningkat disebabkan oleh bobot ubi yang meningkat sejalan dengan umur panen. Oleh karena itu peningkatan hasil pati dapat diperoleh melalui penundaan umur panen dalam upaya meningkatkan hasil pati tiap satuan waktu dan luas lahan, yaitu antara 9,63–17,02 kg/hari/ha.
Bahan campuran premium 10% (premium mix E10) memiliki angka oktan sekitar 10% lebih tinggi dibandingkan dengan premium murni. Penggunaan premium mix E10 tersebut tidak memerlukan perubahan bagian-bagian pada mesin mobil untuk transportasi. Tingginya hasil pati tiap satuan luas dan waktu serta angka oktan tersebut memberikan gambaran bahwa ubi kayu sebagai bahan baku industri bioetanol berpotensi dikembangkan dalam program pemanfaatan bahan bakar nabati (biofuel) untuk bidang transportasi.
Efisiensi penggunaan air tinggi
Ketersediaan air untuk pertumbuhan ubi kayu yang ideal setiap fase pertumbuhan sekitar 30 mm, 50 mm, dan 30 mm/10 hari, masing-masing selama tiga bulan pertama, tiga bulan ke dua, dan satu bulan sebelum panen, Ubi kayu dapat mengatasi cekaman kekurangan air melalui aktivitas akar dan kanopi, di mana pada kondisi air tidak tersedia di dalam tanah lapisan atas, sekitar 30% dari akar ubi kayu berpenetrasi ke lapisan yang lebih bawah sampai kedalaman sekitar tiga meter, dan tanaman menggugurkan seluruh daunnya (kecuali daun pucuk) agar air yang diserap akar tidak menguap melalui helaian daun. Melalui mekanisme tersebut ubi kayu dapat bertahan hidup walaupun terjadi kekeringan. Implikasi dari keunggulan tersebut adalah ubi kayu potensial dikembangkan pada berbagai tipe iklim dan pola tanam, yaitu (1) wilayah beriklim basah dan kering, (2) dapat ditanam pada musim hujan dan kemarau, dan (3) dapat dirotasi dengan tanaman yang memerlukan air relatif tinggi dengan pola rotasi padi gogo, ubi kayu, padi sawah–ubi kayu, jagung–ubi kayu, dan aneka kacang ubi kayu.
Efisiensi penggunaan lahan tinggi
Berdasarkan indikator rasio efisiensi penggunaan lahan (LER) dan potensi energi, ubi kayu mampu menggunakan lahan lebih efisien dibanding tanaman pangan lain. Ubi kayu memiliki pertumbuhan lambat selama tiga bulan pertama dan periode akumulasi fotosintat lama (12 bulan atau lebih), sehingga dapat dimanfaatkan untuk meningkatkan efisiensi penggunaan lahan melalui tumpang sari, seperti ubi kayu+padi/aneka kacang + jagung – aneka kacang-kacangan (jarak tanam ubi kayu 250x50 cm) atau sistem rotasi padi/palawija–ubi kayu. Penggunaan tumpangsari dengan pengelolaan optimal dan pemilihan varietas tanaman sela yang tepat, efisiensi penggunaan lahannya (LER) mencapai 2,5 atau lebih (Wargiono et al. 2001). Ubi kayu juga Prospek, Strategi dan Teknologi Pengembangan Ubi kayu untuk Agroindustri dan Ketahanan Pangan
memiliki efisiensi penggunaan lahan berdasarkan indikator hasil persatuan luas dan waktu paling tinggi dibanding padi sawah, padi gogo dan jagung, baik berdasarkan parameter hasil maupun kalori (Tabel 2). Berdasarkan indikator efisiensi penggunaan lahan tersebut ubi kayu sebagai bahan baku industri bioetanol cukup potensial untuk dikembangkan, baik pada lahan kering maupun lahan sawah tadah hujan Indeks Pertanaman padi 100.
Usaha tani sistem integrasi ubi kayu–ternak
Sebagian besar lahan pertanian di Indonesia miskin bahan organik, sedangkan petani ubi kayu yang menggunakan pupuk organik sesuai dengan kondisi tanah (rasional) dan kontinyu relatif terbatas. Oleh karena penggunaan pupuk organik secara kontinyu sesuai dengan kondisi tanah dapat mempertahankan stabilitas hasil, meningkatkan efisiensi penggunaan pupuk anorganik dan memperbaiki fisik tanah, maka perlu adanya upaya untuk menjamin ketersediaan pupuk organik sesuai dengan kebutuhan dan murah. Salah satu cara untuk mencapai hal tersebut adalah usaha tani sistem integrasi ubi kayu–ternak ruminansia. Limbah ubi kayu yang dapat digunakan sebagai pakan ternak adalah daun dan kulit ubi, baik langsung maupun melalui olahan dalam bentuk silase. Daun ubi kayu berkadar protein tinggi, oleh karena itu penggunaan daun ubi
kayu sebagai suplemen pakan ruminansia (rumput gajah, jerami, dan rumput lainnya) dapat meningkatkan bobot harian ruminansia paling tinggi di- banding penggunaan Gliricedia dan Leucaena (Wargiono 2005). Rumput Gajah dapat diperoleh dari tanaman sistem hedgerows pada pematang yang ditanam baik khusus untuk pakan maupun pengendali erosi. Sedangkan jerami padi dan jagung dapat diperoleh dari tanaman sela dalam sistem tumpangsari atau rotasi. Hasil limbah panen dalam sistem tumpangsari ubi kayu+padi+jagung– kacang tanah dapat menyediakan pakan sekitar 48 ekor ruminansia kecil atau tiga ekor ruminansia besar/ha/tahun. Oleh karena limbah panen dari usaha tani dapat menjamin ketersediaan pakan selama setahun, penggunaan limbah untuk pakan dapat meningkatkan bobot harian serta hasil pupuk organik, dan penggunaan pupuk organik secara kontinyu dapat meningkatkan hasil dan limbah panen, maka usaha tani sistem integrasi ubi kayu–ternak tersebut sinergis, sehingga sistem tersebut potensial dikembangkan dalam upaya meningkatkan produktivitas ubi kayu secara lestari dengan input eksternal minimal (LEISA).
Sumberdaya lahan
Tipe agroekologi yang sesuai untuk pengembangan ubi kayu sebagai bahan baku industri bioetanol adalah lahan kering dan sawah tadah hujan beriklim basah dengan kategori kesesuaian lahan (S1 dan S2 dan S3 ) dengan perbaikan pengelolaan lahan yang optimal. Potensi hasil ubi kayu pada tanah Inseptisol (alkalin) dan Ultisol (masam) antara 25–35 ton/ha, lahan dengan tingkat kesesuaian S2 dan S3 dengan pengelolaan optimal sangat potensial untuk pengembangan ubi kayu.
Luas pertanaman ubi kayu di sentra produksi yang berpotensi ditingkatkan produktivitasnya di daerah beriklim basah dan kering sekitar 0,4 juta ha dan 0,70 juta ha. Luas lahan tidur di daerah beriklim basah dan kering adalah 4,5 juta dan 1,3 juta ha. Untuk lahan sawah tadah hujan di daerah beriklim basah dan kering masing-masing 0,3 juta dan 0,8 juta ha. Lahan tidur dan sawah tadah hujan tersebut cukup potensial untuk perluasan areal tanaman sebagai kebun penyangga yang dikelola oleh pihak industri bioetanol.
Aspek Sosial Ekonomi
Secara sosial ekonomi, pengembangan ubi kayu berpotensi dikembangkan atas dasar respon petani terhadap usaha tani ubi kayu cukup tinggi, budidaya ubi kayu telah dikenal petani secara turun temurun di 36 propinsi, biaya produksi usaha tani ubi kayu lebih murah dibanding tanaman pangan lainnya, sumbangan ubi kayu terhadap pendapatan petani cukup besar, dan sebagai bahan pangan pokok. Fluktuasi produksi antar waktu yang tinggi sebagai akibat dari fluktuasi harga ubi di tingkat petani mengindikasikan bahwa respon petani terhadap usaha tani juga tinggi bila harga ubi memadai. Dengan demikian, adanya pengembangan industri bioetanol yang dapat menciptakan harga ubi yang layak akan mendorong petani untuk mening-
katkan produksi ubi kayu secara regional.
Budidaya ubi kayu yang telah dikenal petani secara turun-temurun merupakan modal dasar yang dapat dimanfaatkan dalam pengembangan ubi kayu melalui intensifikasi. Senjang produktivitas yang cukup besar antar propinsi dan pertumbuhan produktivitas sekitar 2,58%/tahun selama lima tahun terakhir mengindikasikan bahwa petani responsif terhadap teknologi untuk meningkatkan hasil bila didukung harga yang memadai. Ubi kayu sebagai pangan pokok merupakan faktor pendorong terhadap keberlanjutan usaha tani sejalan dengan kebutuhan pangan keluarga. Oleh karena kebutuhan pangan berdasarkan kesetaraan kalori sekitar 1 ton ubi segar/tahun/keluarga dan luas lahan untuk ubi kayu antara 0,25–0,75 ha/kk, maka akan terdapat kelebihan hasil sebagai bahan baku industri bioetanol.
Biaya produksi usaha tani ubi kayu per satuan luas lebih murah 20–40% dibanding padi gogo dan palawija lainnya, sehingga usaha tani ubi kayu merupakan pilihan bagi petani bermodal terbatas. Selanjutnya, mengingat jumlah petani bermodal terbatas jumlahnya sekitar 35% dari total keluarga tani, maka terdapat potensi menggeser sistem usaha taninya dari subsisten ke semi komersial melalui bantuan kredit usaha tani dalam pola kemitraan dengan pihak industri bioetanol.
Hambatan Pengembangan Ubi Kayu
Hambatan dalam pengembangan ubi kayu sebagai bahan baku industri bioetanol dapat dipilah berdasarkan aspek manajemen, biofisik dan sosial ekonomi.
Aspek Manajemen
Hambatan pengembangan ubi kayu yang berkaitan dengan aspek mana- jemen, di antaranya adalah: (1) program Pemda belum komprehensif, (2) dukungan sistem pemasaran lemah, (3) dukungan BKPM kurang realistis, (4) penyuluhan pertanian bias ke komoditas superior, (5) Perpres No.5 dan Inpres No. 1 tahun 2006 belum terimplementasikan di kabupaten, (6) tata ruang pengembangan industri berbahan baku ubi kayu belum berbasis daya dukung lingkungan, (7) koordinasi antar instansi terkait lemah, (8) ubi kayu belum termasuk komoditas unggulan prioritas utama, (9) koordinasi tim teknis dalam penyusunan teknologi anjuran lemah, dan (10) infrastruktur belum memadai.
Program Pemda untuk pengembangan industri berbahan baku ubi kayu kurang komprehensif berdasarkan indikator belum tercerminnya keterkaitan antara daya dukung lingkungan dengan industri hulu dan hilir. Tata ruang pembangunan fisik industri berbahan baku ubi kayu di beberapa propinsi tidak berdasarkan pada daya dukung lingkungan seperti jarak antar industri berdasarkan radius kemampuan hamparan untuk memenuhi bahan baku industri; misalnya, industri dengan kapasitas 390 ton ubi segar/hari mempunyai wilayah pendukung sekitar 30 km untuk wilayah dengan agroekosistem di mana ubi kayu hanya mampu memberikan hasil 20 ton/ha ubi segar.
Peluang investor untuk membangun industri bioetanol sulit mendapatkan persetujuan BKPM karena terkait dengan industri alkohol yang berpeluang mengarah ke minuman keras. Masalah yang cukup pelik adalah adanya cukai untuk produk-produk alkohol. Bila hal tersebut berlaku untuk bioetanol sebagai campuran bahan bakar, maka akan memberatkan pihak industri untuk mendapatkan harga bahan baku yang layak bagi petani.
Pasar lokal untuk produk bioetanol belum jelas keberlanjutannya disebabkan oleh Perda penjabaran Keppres No. 05 dan Inpres No. 1 Tahun 2005 yang belum terimplementasi di daerah-daerah proyeksi pengembangan industri bioetanol. Dengan demikian, diperlukan adanya SK Menteri terkait untuk memberikan kepastian pasar sebagai pegangan bagi investor untuk mengembangkan industri bioetanol.
Posisi Penyuluh Pertanian Lapang (PPL) yang cukup solid pada saat revolusi hijau cenderung melemah dalam era otonomi daerah. Kondisi tersebut diperparah dengan beralih fungsinya PPL ke struktural dan PPL yang ada cenderung bias ke komoditas superior. Bila revitalisasi lembaga penyuluhan tidak dilaksanakan, keberhasilan pengembangan bahan bakar nabati untuk mengurangi ketergantungan terhadap BBM akan terhambat. Keputusan Presiden No. 05 dan Instruksi Presiden No. 1 tahun 2006 belum dilengkapi dengan SK tentang BBM mix, harga dan sistem distribusinya di SPBU belum merupakan kekuatan penuh bagi para investor untuk mengembangkan industri bioetanol. Bila Perda dan SK Menteri terkait tidak segera direalisasikan percepatan pengembangan industri bioetanol akan terhambat.
Tata ruang industri bioetanol yang ideal adalah yang berbasis daya dukung lingkungan yakni dalam bentuk hamparan lahan yang produksi ubi kayunya mampu memenuhi kebutuhan bahan baku harian selama setahun secara kontinyu. Sebagai contoh adalah setiap hamparan pertanaman ubi kayu dengan produktivitas 20 ton/ha dalam radius 30 km hanya boleh dibangun satu industri bioetanol dengan kapasitas 60 kl/hari. Dalam hamparan tersebut termasuk perkebunan energi yang dikelola oleh industri sebagai penyangga ketersediaan bahan baku. Ketentuan tersebut harus
termasuk dalam Perda. Koordinasi antar instansi terkait dengan progaram aksi pengembangan industri bioetanol dari pusat sampai daerah belum terwujud dalam suatu
kesepakatan. Bila hal tersebut tidak segera terimplementasi berpotensi menghambat investor untuk mengembangkan industri bioetanol di daerah sentra produksi ubi kayu.
Produktivitas ubi kayu yang mempunyai keunggulan dibanding komoditas tanaman pangan lain dalam usaha tani dipengaruhi oleh penggunaan teknologi inovatif spesifik lokasi. Teknologi produksi tersebut disusun berdasarkan hasil kajian dari BPTP bekerjasama dengan instansi terkait yang disusun oleh tim teknis. Otonomi daerah yang organisasinya inkonsisten mengakibatkan koordinasi tim teknis juga tidak konsisten. Bila hal tersebut tidak dibenahi berpotensi menghambat partisipasi petani dalam pengem- bangan industri bioetanol.
Petani produsen ubi kayu mengalami kesulitan dalam pemasaran hasil untuk mendapatkan harga yang layak, disebabkan oleh dukungan fasilitas dan infrastruktur yang belum memadai. Implikasinya adalah, industri bioetanol akan mengalami kesulitan untuk mendapatkan bahan baku dengan harga layak pada musim hujan disebabkan oleh sulitnya armada angkutan mencapai lahan petani yang infrastrukturnya belum memadai. Industri tapioka tidak tersebar sesuai dengan sebaran konsumen. Industri tapioka skala besar terkonsentrasi di Sumatera bagian selatan.
Peluang Pasar
konsumen tersebar di berbagai pulau. Kondisi tersebut berpotensi men-
dorong terjadinya harga tapioka impor dari Thailand lebih murah di daerah yang letaknya lebih dekat dengan Thailand dibanding Lampung. Hal tersebut berpeluang terjadi pada industri bioetanol bila sebaran industri bioetanol seperti sebaran industri tapioka, sehingga harga bioetanol akan bervariasi dan tidak kompetitif.
Strategi untuk mengatasi masalah manajemen tersebut diprioritaskan
pada:
• penegakan Perpres/Inpres agar lebih konsisten untuk mendorong minat
swasta mengembangkan industri bioetanol, dan melalui revitalisasi perta-
nian dan perhatian pemerintah yang besar untuk memenuhi permintaan
bioetanol yang tinggi,
• pemanfaatan upaya Pemda menegakkan Perpres No. 5 tahun 2006/Inpres
No. 1 tahun 2006 yang makin konsisten dan pengambilan keputusan melalui
Perda untuk meningkatkan koordinasi antar instansi terkait, dan permintaan
bioetanol yang tinggi untuk memperbaiki implementasi program pengem-
bangan ubi kayu sebagai bahan baku industri bioetanol,
• pemanfaatan perhatian pemerintah yang besar dan minat swasta
mengembangkan industri bioetanol untuk mencegah ekspor gaplek sebagai
bahan baku industri bioetanol skala besar di RRC, dan melalui revitalisasi
pertanian ekspor bioetanol oleh investor asing dan ekspor alkohol dapat
dicegah, dan
• revitalisasi lembaga penyuluhan dan perbaikan koordinasi antar instansi
yang terkait dengan pengembangan ubi kayu sebagai bahan baku industri
bioetanol untuk mencegah ekspor gaplek sebagai bahan baku industri
bioetanol RRC, ekspor bioetanol dan alkohol.
Aspek Biofisik
Hambatan yang terkait dengan aspek biofisik dalam pengembangan ubi kayu untuk memenuhi kebutuhan bahan baku industri bioetanol meliputi sumberdaya tanaman, lahan, dan petani. Permasalahan sumberdaya tanaman meliputi: (1) siklus pertanaman panjang, (2) pertumbuhan fase awal lambat, (3) kadar pati bervariasi dan spesifik agroekologi, (4) sifat hasil “bulky”. Permasalahan sumberdaya lahan meliputi: (1) kadar hara makro dan bahan organik tanah rendah, (2) peka erosi dan konservasinya belum efektif, (3) PTT belum berkembang, (4) sentra produksi didominasi oleh iklim kering, (5) kinerja kelompok tani lemah, (6) tenaga kerja produktif untuk usaha tani terbatas, dan (7) tingkat pendidikan rendah.
Dampak negatif dari umur tanaman yang panjang dalam usaha tani ubi kayu berdasarkan hasil pengamatan lapangan di daerah sentra produksi, diantaranya adalah: (1) investasi input rendah, (2) sering dipanen sebelum umur optimal, (3) sistem usaha tani subsisten, (4) didominasi oleh sistem tumpang sari untuk petani berlahan sempit, (5) sistem rotasi dengan tanaman pangan berumur pendek, dan (6) pengelolaan kurang optimal. Usaha tani ubi kayu dengan sistem tersebut produktivitasnya rendah dan cenderung tidak mempunyai keunggulan. Kondisi tersebut dapat menghambat pemenuhan bahan baku industri bioetanol. Pertumbuhan ubi kayu pada fase awal yang lambat berpotensi memicu munculnya masalah seperti erosi dan gangguan gulma. Erosi akan berlangsung sejalan dengan waktu yang menyebabkan terjadinya degradasi tanah bila tidak dilakukan konservasi yang efektif. Pertumbuhan ubi kayu yang lambat pada fase awal memacu gulma untuk tumbuh lebih cepat dan pertumbuhan ubi kayu terhambat karena tidak mampu berkompetisi dengan gulma. Bila pengendalian erosi dan gulma tidak dilakukan secara efektif, produktivitas ubi kayu tidak optimal sehingga berpotensi tidak terpenuhinya kebutuhan bahan baku untuk industri bioetanol.
Kadar pati maksimal berdasarkan potensi genetis dari varietas unggul dapat dicapai pada kondisi lingkungan optimal, dan akan menurun bila terjadi cekaman lingkungan. Kondisi optimal untuk tiap varietas tidak sama, sehingga penggunaan multivarietas unggul (umur genjah, sedang dan dalam) diperlukan pada daerah yang kondisi lingkungan abiotiknya bervariasi, seperti kemarau panjang atau anomali iklim, dan lain-lain.
Sistem tumpangsari ubi kayu dengan tanaman pangan lainnya berpotensi
menurunkan produktivitasnya bila terjadi kompetisi hara dan cahaya, namun berpeluang meningkatkan produktivitas bila pengaturan jenis tanaman dan populasi tanaman/jarak tanamnya tepat. Oleh karena jumlah petani berlahan sempit mendominasi petani di pulau Jawa, maka kondisi tersebut berpotensi mengganggu ketersediaan bahan baku industri bioetanol.
Sebagian lahan pertanian berkadar hara makro dan bahan organik antara rendah – sangat rendah. Hal tersebut dapat menjadi faktor pembatas produktivitas. Bila kondisi tersebut tidak diatasi, maka dapat menjadi faktor penghambat dalam upaya meningkatkan produksi ubi kayu untuk mendukung pengembangan industri bioetanol. Upaya tersebut sulit diimplementasikan disebabkan terbatasnya modal usaha tani dan petani yang memiliki ruminansia sebagai produsen pupuk organik jumlahnya relatif sedikit.Iklim kering mendominasi daerah sentra produksi ubi kayu. Karakteristik produksi ubi kayu di daerah beriklim kering adalah petaninya menanam ubi kayu secara serentak pada awal musim hujan dan bulan panen terkonsentrasi
pada musim kemarau. Dengan demikian produksi ubi kayu tidak terdistribusi merata sepanjang tahun sesuai dengan kebutuhan bahan baku industri bioetanol.Rataan hasil ubi kayu nasional sekitar 40–60% dari potensi genetis merupakan indikator bahwa pengelolaan tanaman terpadu (PTT) belum berkembang. Lahan marginal dan usaha tani subsisten yang mendominasi sebagian besar daerah sentra produksi ubi kayu merupakan penyebab utama belum berkembangnya teknologi pengelolaan tanaman terpadu/sistem integrasi ubi kayu–ternak (PTT/SIUT).
Kelembagaan Lokal dan SDM
Proyek Kemitraan Terpadu (PKT) adalah suatu program kemitraan terpadu yang melibatkanusaha besar (inti), usaha kecil (plasma) dengan melibatkan bank sebagai pemberi kreditdalam suatu ikatan kerja sama yang dituangkan dalam nota kesepakatan. Tujuan PKTantara lain adalah untuk meningkatkan kelayakan plasma, meningkatkan keterkaitan dankerjasama yang saling menguntungkan antara inti dan plasma, serta membantu bankdalam meningkatkan kredit usaha kecil secara lebih aman dan efisien.Dalam melakukan kemitraan hubunga kemitraan, perusahaan inti (Industri Pengolahan atauEksportir) dan petani plasma/usaha kecil mempunyai kedudukan hukum yang setara.Kemitraan dilaksanakan dengan disertai pembinaan oleh perusahaan inti, dimulai daripenyediaan sarana produksi, bimbingan teknis dan pemasaran hasil produksi.
Proyek Kemitraan Terpadu ini merupakan kerjasama kemitraan dalam bidang usahamelibatkan tiga unsur, yaitu (1) Petani/Kelompok Tani atau usaha kecil, (2) PengusahaBesar atau eksportir, dan (3) Bank pemberi KKPA.Masing-masing pihak memiliki peranan di dalam PKT yang sesuai dengan bidang usahanya.Hubungan kerjasama antara kelompok petani/usaha kecil dengan Pengusaha Pengolahanatau eksportir dalam PKT, dibuat seperti halnya hubungan antara Plasma dengan Inti didalam Pola Perusahaan Inti Rakyat (PIR). Petani/usaha kecil merupakan plasma danPerusahaan Pengelolaan/Eksportir sebagai Inti. Kerjasama kemitraan ini kemudian menjaditerpadu dengan keikut sertaan pihak bank yang memberi bantuan pinjaman bagipembiayaan usaha petani plasma. Proyek ini kemudian dikenal sebagai PKT yang disiapkandengan mendasarkan pada adanya saling berkepentingan diantara semua pihak yang bermitra.
1. Petani Plasma
Sesuai keperluan, petani yang dapat ikut dalam proyek ini bisa terdiri atas (a) Petani yangakan menggunakan lahan usaha pertaniannya untuk penanaman dan perkebunan atauusaha kecil lain, (b) Petani /usaha kecil yang telah memiliki usaha tetapi dalam keadaanyang perlu ditingkatkan dalam untuk itu memerlukan bantuan modal.Untuk kelompok (a), kegiatan proyek dimulai dari penyiapan lahan dan penanaman ataupenyiapan usaha, sedangkan untuk kelompok (b), kegiatan dimulai dari telah adanya kebunatau usaha yang berjalan, dalam batas masih bisa ditingkatkan produktivitasnya denganperbaikan pada aspek usaha.Luas lahan atau skala usaha bisa bervariasi sesuai luasan atau skala yang dimiliki olehmasing-masing petani/usaha kecil. Pada setiap kelompok tani/kelompok usaha, ditunjukseorang Ketua dan Sekretaris merangkap Bendahara. Tugas Ketua dan Sekretaris Kelompokadalah mengadakan koordinasi untuk pelaksanaan kegiatan yang harus dilakukan oleh parapetani anggotanya, didalam mengadakan hubungan dengan pihak Koperasi dan instansilainnya yang perlu, sesuai hasil kesepakatan anggota. Ketua kelompok wajib menyelenggarakan pertemuan kelompok secara rutin yang waktunya ditentukan berdasarkan kesepakatan kelompok.
2. Koperasi
Para petani/usaha kecil plasma sebagai peserta suatu PKT, sebaiknya menjadi anggotasuata koperasi primer di tempatnya. Koperasi bisa melakukan kegiatan-kegiatan untuk membantu plasma di dalam pembangunan kebun/usaha sesuai keperluannya. Fasilitas KKPA hanya bisa diperoleh melalui keanggotaan koperasi. Koperasi yang mengusahakanKKPA harus sudah berbadan hukum dan memiliki kemampuan serta fasilitas yang cukupbaik untuk keperluan pengelolaan administrasi pinjaman KKPA para anggotanya. Jikamenggunakan skim Kredit Usaha Kecil (KUK), kehadiran koperasi primer tidak merupakankeharusan
3. Perusahaan Besar dan Pengelola/Eksportir
Suatu Perusahaan dan Pengelola/Eksportir yang bersedia menjalin kerjasama sebagai intidalam Proyek Kemitraan terpadu ini, harus memiliki kemampuan dan fasilitas pengolahanuntuk bisa menlakukan ekspor, serta bersedia membeli seluruh produksi dari plasma untukselanjutnya diolah di pabrik dan atau diekspor. Disamping ini, perusahaan inti perlumemberikan bimbingan teknis usaha dan membantu dalam pengadaan sarana produksiuntuk keperluan petani plasma/usaha kecil.Apabila Perusahaan Mitra tidak memiliki kemampuan cukup untuk mengadakan pembinaanteknis usaha, PKT tetap akan bisa dikembangkan dengan sekurang-kurangnya pihak Intimemiliki fasilitas pengolahan untuk diekspor, hal ini penting untuk memastikan adanyapemasaran bagi produksi petani atau plasma. Meskipun demikian petani plasma/usaha kecildimungkinkan untuk mengolah hasil panennya, yang kemudian harus dijual kepadaPerusahaan Inti.Dalam hal perusahaan inti tidak bisa melakukan pembinaan teknis, kegiatan pembibinganharus dapat diadakan oleh Koperasi dengan memanfaatkan bantuan tenaga pihak DinasPerkebunan atau lainnya yang dikoordinasikan oleh Koperasi. Apabila koperasimenggunakan tenaga Penyuluh Pertanian Lapangan (PPL), perlu mendapatkan persetujuanDinas Perkebunan setempat dan koperasi memberikan bantuan biaya yang diperlukan.Koperasi juga bisa memperkerjakan langsung tenaga-tenaga teknis yang memilikiketerampilan dibidang perkebunan/usaha untuk membimbing petani/usaha kecil dengandibiayai sendiri oleh Koperasi. Tenaga-tenaga ini bisa diberi honorarium oleh Koperasi yangbisa kemudian dibebankan kepada petani, dari hasil penjualan secara proposional menurutbesarnya produksi. Sehingga makin tinggi produksi kebun petani/usaha kecil, akan semakinbesar pula honor yang diterimanya.
4. Bank
Bank berdasarkan adanya kelayakan usaha dalam kemitraan antara pihak Petani Plasmadengan Perusahaan Perkebunan dan Pengolahan/Eksportir sebagai inti, dapat kemudianmelibatkan diri untuk biaya investasi dan modal kerja pembangunan atau perbaikan kebun.Disamping mengadakan pengamatan terhadap kelayakan aspek-aspek budidaya/produksiyang diperlukan, termasuk kelayakan keuangan. Pihak bank di dalam mengadakan evaluasi, juga harus memastikan bagaimana pengelolaan kredit dan persyaratan lainnya yang diperlukan sehingga dapat menunjang keberhasilan proyek. Skim kredit yang akandigunakan untuk pembiayaan ini, bisa dipilih berdasarkan besarnya tingkat bunga yangsesuai dengan bentuk usaha tani ini, sehingga mengarah pada perolehannya pendapatanbersih petani yang paling besar.Dalam pelaksanaanya, Bank harus dapat mengatur cara petani plasma akan mencairkankredit dan mempergunakannya untuk keperluan operasional lapangan, dan bagaimanapetani akan membayar angsuran pengembalian pokok pinjaman beserta bunganya. Untukini, bank agar membuat perjanjian kerjasama dengan pihak perusahaan inti, berdasarkankesepakatan pihak petani/kelompok tani/koperasi. Perusahaan inti akan memotong uanghasil penjualan petani plasma/usaha kecil sejumlah yang disepakati bersama untukdibayarkan langsung kepada bank. Besarnya potongan disesuaikan dengan rencanaangsuran yang telah dibuat pada waktu perjanjian kredit dibuat oleh pihak petani/Kelompoktani/koperasi. Perusahaan inti akan memotong uang hasil penjualan petani plasma/usahakecil sejumlah yang disepakati bersama untuk dibayarkan langsung kepada Bank. Besarnyapotongan disesuaikan dengan rencana angsuran yang telah dibuat pada waktu perjanjiankredit dibuat oleh pihak petani plasma dengan bank.
Pola Kerjasama
Kemitraan antara petani/kelompok tani/koperasi dengan perusahaan mitra, dapat dibuatmenurut dua pola yaitu :a. Petani yang tergabung dalam kelompok-kelompok tani mengadakan perjanjian kerjasamalangsung kepada Perusahaan Perkebunan/Pengolahan Eksportir.Dengan bentuk kerja sama seperti ini, pemberian kredit yang berupa KKPA kepada petaniplasma dilakukan dengan kedudukan Koperasi sebagai Channeling Agent, danpengelolaannya langsung ditangani oleh Kelompok tani. Sedangkan masalah pembinaanharus bisa diberikan oleh Perusahaan Mitra.b. Petani yang tergabung dalam kelompok-kelompok tani, melalui koperasinya mengadakanperjanjian yang dibuat antara Koperasi (mewakili anggotanya) dengan perusahaanperkebunan/pengolahan/eksportir.
Petani yang tergabung dalam kelompok-kelompok tani mengadakan perjanjian kerjasamalangsung kepada Perusahaan Perkebunan/Pengolahan Eksportir.Dengan bentuk kerja sama seperti ini, pemberian kredit yang berupa KKPA kepada petaniplasma dilakukan dengan kedudukan Koperasi sebagai Channeling Agent, danpengelolaannya langsung ditangani oleh Kelompok tani. Sedangkan masalah pembinaanharus bisa diberikan oleh Perusahaan Mitra.b. Petani yang tergabung dalam kelompok - kelompok tani, melalui koperasinya mengadakan perjanjian yang dibuat antara Koperasi (mewakili anggotanya) dengan perusahaanperkebunan/pengolahan/eksportir.
Dalam bentuk kerjasama seperti ini, pemberian KKPA kepada petani plasma dilakukandengan kedudukan koperasi sebagai Executing Agent. Masalah pembinaan teknis budidayatanaman/pengelolaan usaha, apabila tidak dapat dilaksanakan oleh pihak Perusahaan Mitra,akan menjadi tanggung jawab koperasi.
Penyiapan Proyek Kemitraan Terpadu
Untuk melihat bahwa PKT ini dikembangkan dengan sebaiknya dan dalam proseskegiatannya nanti memperoleh kelancaran dan keberhasilan, minimal dapat dilihat daribagaimana PKT ini disiapkan. Kalau PKT ini akan mempergunakan KKPA untuk modal usahaplasma, perintisannya dimulai dari
A. Adanya petani/pengusaha kecil yang telah menjadi anggota koperasi dan lahan pemilikannya akan dijadikan kebun/tempat usaha atau lahan kebun/usahanya sudah adatetapi akan ditingkatkan produktivitasnya. Petani/usaha kecil tersebut harus menghimpundiri dalam kelompok dengan anggota sekitar 25 petani/kelompok usaha. Berdasarkan persetujuan bersama, yang didapatkan melalui pertemuan anggota kelompok, merekabersedia atau berkeinginan untuk bekerja sama dengan perusahaanperkebunan/pengolahan/eksportir dan bersedia mengajukan permohonan kredit (KKPA)untuk keperluan peningkatan usaha. Adanya perusahaan perkebunan/pengolahan dan eksportir, yang bersedia menjadimitra petani/usaha kecil, dan dapat membantu memberikan pembinaan teknikbudidaya/produksi serta proses pemasarannya, Dipertemukannya kelompok tani/usaha kecil dan pengusaha perkebunan/pengolahandan eksportir tersebut, untuk memperoleh kesepakatan di antara keduanya untuk bermitra.Prakarsa bisa dimulai dari salah satu pihak untuk mengadakan pendekatan, atau ada pihakyang akan membantu sebagai mediator, peran konsultan bisa dimanfaatkan untukmengadakan identifikasi dan menghubungkan pihak kelompok tani/usaha kecil yangpotensial dengan perusahaan yang dipilih memiliki kemampuan tinggi memberikan fasilitasyang diperlukan oleh pihak petani/usaha kecil;
Diperoleh dukungan untuk kemitraan yang melibatkan para anggotanya oleh pihakkoperasi. Koperasi harus memiliki kemampuan di dalam mengorganisasikan dan mengelolaadministrasi yang berkaitan dengan PKT ini. Apabila keterampilan koperasi kurang, untukpeningkatannya dapat diharapkan nantinya mendapat pembinaan dari perusahaan mitra.Koperasi kemudian mengadakan langkah-langkah yang berkaitan dengan formalitas PKTsesuai fungsinya. Dalam kaitannya dengan penggunaan KKPA, Koperasi harus mendapatkanpersetujuan dari para anggotanya, apakah akan beritndak sebagai badan pelaksana(executing agent) atau badan penyalur (channeling agent);
Diperolehnya rekomendasi tentang pengembangan PKT ini oleh pihak instansipemerintah setempat yang berkaitan (Dinas Perkebunan, Dinas Koperasi, KantorBadan Pertanahan, dan Pemda);
Mekanisme Proyek Kemitraan Terpadu
Mekanisme Proyek Kemitraan Terpadu dapat dilihat pada skema berikut ini Bank pelaksana akan menilai kelayakan usaha sesuai dengan prinsip-prinsip bank teknis.Jika proyek layak untuk dikembangkan, perlu dibuat suatu nota kesepakatan (Memorandumof Understanding = MoU) yang mengikat hak dan kewajiban masing-masing pihak yang bermitra (inti, Plasma/Koperasi dan Bank). Sesuai dengan nota kesepakatan, atas kuasakoperasi atau plasma, kredit perbankan dapat dialihkan dari rekening koperasi/plasma kerekening inti untuk selanjutnya disalurkan ke plasma dalam bentuk sarana produksi, danapekerjaan fisik, dan lain-lain.
Dengan demikian plasma tidak akan menerima uang tunai dariperbankan, tetapi yang diterima adalah sarana produksi pertanian yang penyalurannyadapat melalui inti atau koperasi. Petani plasma melaksanakan proses produksi. Hasiltanaman plasma dijual ke inti dengan harga yang telah disepakati dalam MoU. Perusahaaninti akan memotong sebagian hasil penjualan plasma untuk diserahkan kepada banksebagai angsuran pinjaman dan sisanya dikembalikan ke petani sebagai pendapatan bersih.
DAFTAR PUSTAKA
Anonim, 2000. Ketika Kebun Berupa Hutan. Agroforestri Khas Indonesia.
Sebuah Sumbangan Masyarakat. International Centre For Research In
Agroforestry. Bogor.
Anonim. 2001. Sistem Pertanian di Indonesia.http://www.lab link.or.id. Diakses
pada tanggal 22 Mei 2010 pukul 17.00 WIB
Danoesastro, Haryono. 1979. Pemanfaatan Pekarangan. Yayaan Pembina FakulasPertanian UGM. Yogyakarta
.
Dover,M. dan Talbot,L.M., 1987. To Feed The Earth: Agroecology for
Sustainable Development. World Resources Intitute. Washington DC.
Handayanto, E. 1999. Pengelolan Kesuburan Tanah. Fakultas Pertanian.
Universitas Brawijaya. Malang.
Hardjowigeno, S., 1989. Ilmu Tanah. Mediyatama Sarana Perkasa, Jakarta
Manuwoto.2009. Sistem Pertanian di Indonesia.
Http://makhey.blogspot.com/2009/09/sistem-pertanian-di-indonesia.
Diakses pada tanggal 27 Mei 2010.
Monika, WT et al. 1993. Produksi Kambing dan Domba di Indonesia. Sebelas
Maret Universitas Press. Surakarta.
Notohadinagoro, Tejoyuwono. 1997. Bercari manat Pengelolaan Berkelanjutan
Sebagai Konsep Pengembangan Wilayah Lahan Kering.Makalah
Seminar Nasional dan Pelatihan Pengelolaan Lahan Kering FOKUSHIMITI di Jember. Universitas Jember. Jember
Pusat Peneliti Universitas Brawijaya. 1991. Penelitian dan Pengembangan Sistem
Usaha Tani Lahan Kering Yang Berkelanjutan. Proseding Simposium Nasional Malang. Universitas Brawijaya. Malang
Reijntjes,C., B.Haverkot dan A. W. Bayer., 1999. Pertanian Masa Depan Pengantar untuk Pertanian Berkelanjutan Dengan Input Luar Rendah. Kanisius. Yogyakarta.
TUGAS KELOMPOK
SISTEM PERTANIAN TERPADU
EKOSISTEM TANAMAN UBI KAYU
DISUSUN OLEH :
ERNAWATY DJAYA ( G111 09 276)
RATI PRADHANI ( G111 09 294 )
NUR JANNAH
IRAWAN TAMRIN
PROGRAM STUDI AGROTEKNOLOGI
JURUSAN ILMU HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
UNIVERSITAS HASANUDDIN
MAKASSAR
2011
Tanaman ubi kayu (Manihotutikisima) memiliki berbagai varietas atau klon yang dapat langsung dikonsumsi sebagai makanan atau menjadi bahan baku bagi industri tapioka dan gapiek (manihok) ataupun tepung gapiek, yang selanjutnya dipergunakan untuk berbagai macam industri seperti makanan, makanan ternak, kertas, kayu lapis dan tainnya. Berdasarkan potensi fisik seperti kesesuaian lahan, iklim, sumberdaya manusia,dan tingkat adaptasi teknologi, tanaman ubi kayu banyak didapatkan dan bisa dibudidayakan di banyak tempat atau lokasi di Indonesia sehingga memungkinkan untuk diusahakan oleh para petani secara luas.
Hasil olahan ubi kayu berupa tapioka dan gapiek (manihok) dalam bentuk chips, pellet ataupun lainnya, telah lama menjadi komoditi ekspor yang sangat penting dalam menyumbang pendapatan devisa, karenanya merupakan aset yang sangat berharga dan perlu dijaga kelestariannya sehingga dapat dimanfaatkan untuk pengembangan ekspor pada masa-masa selanjutnya. Pengalaman petani menunjukkan bahwa penanaman ubi kayu sering tidak membuahkan hasil yang cukup baik untuk keluarga, karena keadaan tata niaga kayu yang banyak dipengaruhi oleh fluktuasi harga sehingga merugikan petani. Pada saat menjelang tanam, harga ubi kayu biasanya terlihat sangat menarik sehingga banyak petani berusaha menanamnya. Akan tetapi pada saat panen harga kayu kemudian jatuh sehingga banyak merugikan petani (berdasarkan data yang ada kapasitas pabrik dan potensi ekspor masih lebih besar dari jumlah produksi).
Kondisi ini telah mendorong banyak petani untuk mengalihkan perhatian dan berusaha menanam komoditi seperti kelapa sawit. Di sisi lain, apabila hal ini terus dibiarkan, akan bisa berakibat turunnya produksi yang dapat menekan pasokan ubi kayu baik untuk keperluan ekspor ataupun konsumsi sawit dalam negeri. Lemahnya posisi petani ubi kayu dalam menghadapi pengaruh fluktuasi harga, terutama disebabkan karena ubi kayu memiliki daya simpan yang rendah,dan produktifitasnya juga rendah akibat modal usaha yang sangat terbatas, disamping kebutuhan keluarga yang sudah sangat mendesak. Pendapatan petani ubi kayu akan makin rendah lagi Karen apada saat dijual ke pabrik mendapatkan mutu ubi kayunya rendah dan rafaksi yang ditentukan secara sepihak oleh pabrik.
Berdasarkan keadaan yang ada, maka salah satu alternatif untuk meng antisipasi masalah tersebut di atas, adalah dengan upaya mengadakan kerjasama antara para petani produsen ubi kayu di satu pihak dengan pengguna atau pemakai ubi kayu (baik dengan kapasitas pabrik yang cukup besar, maupun dengan kapasitas pabrik yang sedang) dipihak lain, sehingga dapat diciptakan bentuk kerjasama kemitraan dengan tata niaga yang saling menguntungkan bagi kedua belah pihak. Untuk mendukung kelancaran kemitraan ini, para petani memerlukan dukungan permodalan untuk keperluan penanaman ubi kayu, di mana bank pelaksana dapat berperan dengan memberikan kredit dengan skim kredit yang sesuai.Secara teknis upaya mengembangkan budidaya ubi kayu harus diikuti dengan adanya upaya untuk mengubah atau memperbaiki teknik budidaya yang selama ini masih konvensional menjadi teknik budidaya yang intensif. Hal ini dapat dicapai dengan lebih memantapkan penataan teknologi produksi mulai dari persiapan lahan, pengolahan lahan, penggunaan varietas unggul, pemeliharaan tanaman, Panen dan pasca panen serta distribusi pemasaran hasil.
TINJAUAN PUSTAKA
Identifikasi Potensi Sumber Daya Alam Tanaman Ubi Kayu
Ubi kayu termasuk tanaman tropis, tetapi dapat pula beradaptasi dan tumbuh dengan baik di daerah subtropis. Secara umum tanaman ini tidak menuntut iklim yang spesifik untuk pertumbuhannya. Namun demikian ubi kayu akan tumbuh dengan baik pada iklim dan tanah. iklim yang cocok untuk tanaman ubi kayu antara lain :
Curah hujan : 750 - 1.000 mm/thn
Tinggi tempat : 0 - 1.500 m dpl
Suhu : 25 ͦ - 28 ͦ C
Fungsi singkong (ubi kayu) sudah mulai bergeser, dari penyediaan bahan pangan, berpotensi menjadi bahan baku untuk pengembangan bio-ethanol. Kebutuhan bio-ethanol sampai dengan 2010 tergolong cukup tinggi, yaitu mencapai 1,8 juta kilo liter.
Pati ataun Amilum
Pati atau amilum adalah karbohidrat kompleks yang tidak larut dalam air, berwujud bubuk putih, tawar dan tidak berbau. Pati merupakan bahan utama yang dihasilkan oleh tumbuhan untuk menyimpan kelebihan glukosa (sebagai produk fotosintesis) dalam jangka panjang. Hewan dan manusia juga menjadikan pati sebagai sumber energi yang penting.
Pati tersusun dari dua macam karbohidrat, amilosa dan amilopektin, dalam komposisi yang berbeda-beda. Amilosa memberikan sifat keras (pera) sedangkan amilopektin menyebabkan sifat lengket. Amilosa memberikan warna ungu pekat pada tes iodin sedangkan amilopektin tidak bereaksi.
“Pati” (bahasa Inggris starch) adalah penyusun (utama) tepung. Tepung bisa jadi tidak murni hanya mengandung pati, karena tercampur dengan protein, pengawet, dan sebagainya. Tepung beras mengandung pati beras, protein, vitamin, dan lain-lain bahan yang terkandung pada butir beras. Orang bisa juga mendapatkan tepung yang merupakan campuran dua atau lebih pati. Kata ‘tepung lebih berkaitan dengan komoditas ekonomis. Pati digunakan sebagai bahan yang digunakan untuk memekatkan makanan cair seperti sup dan sebagainya. Dalam industri, pati dipakai sebagai komponen perekat, campuran kertas dan tekstil, dan pada industri kosmetik.
Bioetanol
Bioetanol merupakan bahan bakar dari minyak nabati yang memiliki sifat menyerupai minyak premium. Untuk pengganti premium, terdapat alternatif gasohol yang merupakan campuran antara bensin dan bioetanol. Adapun manfaat pemakaian gasohol di Indonesia yaitu memperbesar basis sumber daya bahan bakar cair, mengurangi impor BBM, menguatkan security of supply bahan bakar, meningkatkan kesempatan kerja, berpotensi mengurangi ketimpangan pendapatan antar individu dan antar daerah, meningkatkan kemampuan nasional dalam teknologi pertanian dan industri, mengurangi kecenderungan pemanasan global dan pencemaran udara (bahan bakar ramah lingkungan) dan berpotensi mendorong ekspor komoditi baru. Bioetanol tersebut bersumber dari karbohidrat yang potensial sebagai bahan baku seperti jagung, ubi kayu, ubi jalar, sagu dan tebu. Adapun konversi biomasa tanaman tersebut menjadi bioethanol adalah seperti pada tabel dibawah ini.
Tabel Konversi biomasa menjadi bioethanol
Biomassa Jumlah biomassa (kg) Kandungan gula (kg) Jumlah hasil bioetanol (liter) Biomassa : Bioetanol
Ubi Kayu 1.000 250-300 166,6 6,5 : 1
Ubi Jalar 1.000 150-200 125 8 : 1
Jagung 1.000 600-700 400 2,5 : 1
Sagu 1.000 120-160 90 12:1
Tetes 1.000 500 250 4:1
Sumber Daya Tanaman
Potensi pengembangan ubi kayu sebagai bahan bakar industri bioetanol
berdasarkan aspek biofisik di antaranya :
Produktivitas
Rataan produktivitas ubi kayu secara nasional selama dasawarsa terakhir masih rendah, yaitu sekitar 40% dari potensi genetis dengan pengelolaan optimal (BPS 2005; Howeler 2001). Faktor utama yang menyebabkan potensi hasil tinggi adalah kemampuan menghasilkan fotosintat tinggi dan periode pertumbuhannya lama. Tanaman dapat memanfaatkan sinar matahari secara maksimal melalui pengaturan deklinasi dan inklinasi tangkai daun dan helaian daun (Hozyo et al. 1984), di mana helaian daun dapat bergerak mengikuti posisi matahari, sejak pagi hingga sore, ataupun mengikuti pergerakan matahari dari Lintang Selatan ke Lintang Utara dan sebaliknya. Potensi hasil tiap varietas bersifat spesifik lokasi (Suhartina 2005). Oleh karena itu, varietas unggul seperti Adira-4, Malang-6, UJ-3, dan UJ-5 wilayah berkembangnya berbeda. Peningkatan produksi dapat direalisasikan melalui penggunaan varietas unggul dan pengelolaan optimal. Dengan demikian, pengembangan industri pada berbagai tipe agroekologi, ketersediaan bakunya berpotensi dapat dijamin.
Fleksibilitas umur panen
Hasil yang diharapkan dalam pengembangan ubi kayu sebagai bahan baku industri bioetanol adalah ubi berkadar pati tinggi untuk meningkatkan efisiensi industri. Oleh karena itu, kriteria yang harus dipenuhi dalam penentuan umur panen adalah kadar pati optimal. Kadar pati optimal dicapai pada umur yang bervariasi dan secara umum dapat dibedakan menjadi tiga kelompok, yaitu umur: (a) tujuh bulan (genjah), (b) sembilan bulan (sedang), dan (c) 10 bulan (dalam). Kualitas pati tidak berubah walaupun panennya ditunda, sedangkan bobot ubi meningkat sejalan dengan umur. Dengan demikian umur panen dapat disesuaikan dengan kebutuhan industri tanpa penurunan kualitas pati. Kondisi umur panen yang fleksibel tersebut memberikan gambaran adanya potensi untuk menjamin ketersediaan bahan baku industri, sehingga potensial dikembangkan.
Fleksibel dalam usaha tani
Ubi sebagai hasil usaha tani mempunyai peran multi guna, seperti sebagai bahan pangan, industri, pakan dan komoditas ekspor. Oleh karena itu, usaha tani ubi kayu dapat dilakukan secara subsisten, semi komersial dan komersial. Usaha tani subsisten dilakukan petani dengan tujuan utama pemenuhan kebutuhan pangan, sedangkan usaha tani semi komersial dan komersial dengan tujuan utama mendapatkan keuntungan dan pemenuhan permintaan pasar. Ubi kayu pertumbannya lambat selama tiga bulan pertama, sehingga usaha tani sistem tumpangsari dengan padi dan palawija lainnya juga potensial dikembangkan. Ubi kayu dapat dipanen mulai umur tujuh sampai 11 bulan, sehingga dapat dikategorikan sebagai tanaman semusim, dan dapat dipanen pada umur 12 bulan atau lebih, sehingga dapat dikategorikan sebagai tanaman tahunan/tanaman perkebunan. Sifat-sifat tersebut dapat digunakan sebagai indikator bahwa usaha tani ubi kayu fleksibel, dan dalam pengembangannya berpotensi menjamin ketersediaan bahan baku industri bioetanol, pangan, pakan, dan ekspor.
Efisiensi energi tinggi
Sumber energi untuk bioetanol yang berbahan baku ubi kayu adalah pati. Kadar pati varietas unggul yang tersedia berkisar 25–31%. Kadar pati tidak menurun kualitasnya walaupun panennya dilakukan setelah umur optimal, bahkan hasilnya meningkat disebabkan oleh bobot ubi yang meningkat sejalan dengan umur panen. Oleh karena itu peningkatan hasil pati dapat diperoleh melalui penundaan umur panen dalam upaya meningkatkan hasil pati tiap satuan waktu dan luas lahan, yaitu antara 9,63–17,02 kg/hari/ha.
Bahan campuran premium 10% (premium mix E10) memiliki angka oktan sekitar 10% lebih tinggi dibandingkan dengan premium murni. Penggunaan premium mix E10 tersebut tidak memerlukan perubahan bagian-bagian pada mesin mobil untuk transportasi. Tingginya hasil pati tiap satuan luas dan waktu serta angka oktan tersebut memberikan gambaran bahwa ubi kayu sebagai bahan baku industri bioetanol berpotensi dikembangkan dalam program pemanfaatan bahan bakar nabati (biofuel) untuk bidang transportasi.
Efisiensi penggunaan air tinggi
Ketersediaan air untuk pertumbuhan ubi kayu yang ideal setiap fase pertumbuhan sekitar 30 mm, 50 mm, dan 30 mm/10 hari, masing-masing selama tiga bulan pertama, tiga bulan ke dua, dan satu bulan sebelum panen, Ubi kayu dapat mengatasi cekaman kekurangan air melalui aktivitas akar dan kanopi, di mana pada kondisi air tidak tersedia di dalam tanah lapisan atas, sekitar 30% dari akar ubi kayu berpenetrasi ke lapisan yang lebih bawah sampai kedalaman sekitar tiga meter, dan tanaman menggugurkan seluruh daunnya (kecuali daun pucuk) agar air yang diserap akar tidak menguap melalui helaian daun. Melalui mekanisme tersebut ubi kayu dapat bertahan hidup walaupun terjadi kekeringan. Implikasi dari keunggulan tersebut adalah ubi kayu potensial dikembangkan pada berbagai tipe iklim dan pola tanam, yaitu (1) wilayah beriklim basah dan kering, (2) dapat ditanam pada musim hujan dan kemarau, dan (3) dapat dirotasi dengan tanaman yang memerlukan air relatif tinggi dengan pola rotasi padi gogo, ubi kayu, padi sawah–ubi kayu, jagung–ubi kayu, dan aneka kacang ubi kayu.
Efisiensi penggunaan lahan tinggi
Berdasarkan indikator rasio efisiensi penggunaan lahan (LER) dan potensi energi, ubi kayu mampu menggunakan lahan lebih efisien dibanding tanaman pangan lain. Ubi kayu memiliki pertumbuhan lambat selama tiga bulan pertama dan periode akumulasi fotosintat lama (12 bulan atau lebih), sehingga dapat dimanfaatkan untuk meningkatkan efisiensi penggunaan lahan melalui tumpang sari, seperti ubi kayu+padi/aneka kacang + jagung – aneka kacang-kacangan (jarak tanam ubi kayu 250x50 cm) atau sistem rotasi padi/palawija–ubi kayu. Penggunaan tumpangsari dengan pengelolaan optimal dan pemilihan varietas tanaman sela yang tepat, efisiensi penggunaan lahannya (LER) mencapai 2,5 atau lebih (Wargiono et al. 2001). Ubi kayu juga Prospek, Strategi dan Teknologi Pengembangan Ubi kayu untuk Agroindustri dan Ketahanan Pangan
memiliki efisiensi penggunaan lahan berdasarkan indikator hasil persatuan luas dan waktu paling tinggi dibanding padi sawah, padi gogo dan jagung, baik berdasarkan parameter hasil maupun kalori (Tabel 2). Berdasarkan indikator efisiensi penggunaan lahan tersebut ubi kayu sebagai bahan baku industri bioetanol cukup potensial untuk dikembangkan, baik pada lahan kering maupun lahan sawah tadah hujan Indeks Pertanaman padi 100.
Usaha tani sistem integrasi ubi kayu–ternak
Sebagian besar lahan pertanian di Indonesia miskin bahan organik, sedangkan petani ubi kayu yang menggunakan pupuk organik sesuai dengan kondisi tanah (rasional) dan kontinyu relatif terbatas. Oleh karena penggunaan pupuk organik secara kontinyu sesuai dengan kondisi tanah dapat mempertahankan stabilitas hasil, meningkatkan efisiensi penggunaan pupuk anorganik dan memperbaiki fisik tanah, maka perlu adanya upaya untuk menjamin ketersediaan pupuk organik sesuai dengan kebutuhan dan murah. Salah satu cara untuk mencapai hal tersebut adalah usaha tani sistem integrasi ubi kayu–ternak ruminansia. Limbah ubi kayu yang dapat digunakan sebagai pakan ternak adalah daun dan kulit ubi, baik langsung maupun melalui olahan dalam bentuk silase. Daun ubi kayu berkadar protein tinggi, oleh karena itu penggunaan daun ubi
kayu sebagai suplemen pakan ruminansia (rumput gajah, jerami, dan rumput lainnya) dapat meningkatkan bobot harian ruminansia paling tinggi di- banding penggunaan Gliricedia dan Leucaena (Wargiono 2005). Rumput Gajah dapat diperoleh dari tanaman sistem hedgerows pada pematang yang ditanam baik khusus untuk pakan maupun pengendali erosi. Sedangkan jerami padi dan jagung dapat diperoleh dari tanaman sela dalam sistem tumpangsari atau rotasi. Hasil limbah panen dalam sistem tumpangsari ubi kayu+padi+jagung– kacang tanah dapat menyediakan pakan sekitar 48 ekor ruminansia kecil atau tiga ekor ruminansia besar/ha/tahun. Oleh karena limbah panen dari usaha tani dapat menjamin ketersediaan pakan selama setahun, penggunaan limbah untuk pakan dapat meningkatkan bobot harian serta hasil pupuk organik, dan penggunaan pupuk organik secara kontinyu dapat meningkatkan hasil dan limbah panen, maka usaha tani sistem integrasi ubi kayu–ternak tersebut sinergis, sehingga sistem tersebut potensial dikembangkan dalam upaya meningkatkan produktivitas ubi kayu secara lestari dengan input eksternal minimal (LEISA).
Sumberdaya lahan
Tipe agroekologi yang sesuai untuk pengembangan ubi kayu sebagai bahan baku industri bioetanol adalah lahan kering dan sawah tadah hujan beriklim basah dengan kategori kesesuaian lahan (S1 dan S2 dan S3 ) dengan perbaikan pengelolaan lahan yang optimal. Potensi hasil ubi kayu pada tanah Inseptisol (alkalin) dan Ultisol (masam) antara 25–35 ton/ha, lahan dengan tingkat kesesuaian S2 dan S3 dengan pengelolaan optimal sangat potensial untuk pengembangan ubi kayu.
Luas pertanaman ubi kayu di sentra produksi yang berpotensi ditingkatkan produktivitasnya di daerah beriklim basah dan kering sekitar 0,4 juta ha dan 0,70 juta ha. Luas lahan tidur di daerah beriklim basah dan kering adalah 4,5 juta dan 1,3 juta ha. Untuk lahan sawah tadah hujan di daerah beriklim basah dan kering masing-masing 0,3 juta dan 0,8 juta ha. Lahan tidur dan sawah tadah hujan tersebut cukup potensial untuk perluasan areal tanaman sebagai kebun penyangga yang dikelola oleh pihak industri bioetanol.
Aspek Sosial Ekonomi
Secara sosial ekonomi, pengembangan ubi kayu berpotensi dikembangkan atas dasar respon petani terhadap usaha tani ubi kayu cukup tinggi, budidaya ubi kayu telah dikenal petani secara turun temurun di 36 propinsi, biaya produksi usaha tani ubi kayu lebih murah dibanding tanaman pangan lainnya, sumbangan ubi kayu terhadap pendapatan petani cukup besar, dan sebagai bahan pangan pokok. Fluktuasi produksi antar waktu yang tinggi sebagai akibat dari fluktuasi harga ubi di tingkat petani mengindikasikan bahwa respon petani terhadap usaha tani juga tinggi bila harga ubi memadai. Dengan demikian, adanya pengembangan industri bioetanol yang dapat menciptakan harga ubi yang layak akan mendorong petani untuk mening-
katkan produksi ubi kayu secara regional.
Budidaya ubi kayu yang telah dikenal petani secara turun-temurun merupakan modal dasar yang dapat dimanfaatkan dalam pengembangan ubi kayu melalui intensifikasi. Senjang produktivitas yang cukup besar antar propinsi dan pertumbuhan produktivitas sekitar 2,58%/tahun selama lima tahun terakhir mengindikasikan bahwa petani responsif terhadap teknologi untuk meningkatkan hasil bila didukung harga yang memadai. Ubi kayu sebagai pangan pokok merupakan faktor pendorong terhadap keberlanjutan usaha tani sejalan dengan kebutuhan pangan keluarga. Oleh karena kebutuhan pangan berdasarkan kesetaraan kalori sekitar 1 ton ubi segar/tahun/keluarga dan luas lahan untuk ubi kayu antara 0,25–0,75 ha/kk, maka akan terdapat kelebihan hasil sebagai bahan baku industri bioetanol.
Biaya produksi usaha tani ubi kayu per satuan luas lebih murah 20–40% dibanding padi gogo dan palawija lainnya, sehingga usaha tani ubi kayu merupakan pilihan bagi petani bermodal terbatas. Selanjutnya, mengingat jumlah petani bermodal terbatas jumlahnya sekitar 35% dari total keluarga tani, maka terdapat potensi menggeser sistem usaha taninya dari subsisten ke semi komersial melalui bantuan kredit usaha tani dalam pola kemitraan dengan pihak industri bioetanol.
Hambatan Pengembangan Ubi Kayu
Hambatan dalam pengembangan ubi kayu sebagai bahan baku industri bioetanol dapat dipilah berdasarkan aspek manajemen, biofisik dan sosial ekonomi.
Aspek Manajemen
Hambatan pengembangan ubi kayu yang berkaitan dengan aspek mana- jemen, di antaranya adalah: (1) program Pemda belum komprehensif, (2) dukungan sistem pemasaran lemah, (3) dukungan BKPM kurang realistis, (4) penyuluhan pertanian bias ke komoditas superior, (5) Perpres No.5 dan Inpres No. 1 tahun 2006 belum terimplementasikan di kabupaten, (6) tata ruang pengembangan industri berbahan baku ubi kayu belum berbasis daya dukung lingkungan, (7) koordinasi antar instansi terkait lemah, (8) ubi kayu belum termasuk komoditas unggulan prioritas utama, (9) koordinasi tim teknis dalam penyusunan teknologi anjuran lemah, dan (10) infrastruktur belum memadai.
Program Pemda untuk pengembangan industri berbahan baku ubi kayu kurang komprehensif berdasarkan indikator belum tercerminnya keterkaitan antara daya dukung lingkungan dengan industri hulu dan hilir. Tata ruang pembangunan fisik industri berbahan baku ubi kayu di beberapa propinsi tidak berdasarkan pada daya dukung lingkungan seperti jarak antar industri berdasarkan radius kemampuan hamparan untuk memenuhi bahan baku industri; misalnya, industri dengan kapasitas 390 ton ubi segar/hari mempunyai wilayah pendukung sekitar 30 km untuk wilayah dengan agroekosistem di mana ubi kayu hanya mampu memberikan hasil 20 ton/ha ubi segar.
Peluang investor untuk membangun industri bioetanol sulit mendapatkan persetujuan BKPM karena terkait dengan industri alkohol yang berpeluang mengarah ke minuman keras. Masalah yang cukup pelik adalah adanya cukai untuk produk-produk alkohol. Bila hal tersebut berlaku untuk bioetanol sebagai campuran bahan bakar, maka akan memberatkan pihak industri untuk mendapatkan harga bahan baku yang layak bagi petani.
Pasar lokal untuk produk bioetanol belum jelas keberlanjutannya disebabkan oleh Perda penjabaran Keppres No. 05 dan Inpres No. 1 Tahun 2005 yang belum terimplementasi di daerah-daerah proyeksi pengembangan industri bioetanol. Dengan demikian, diperlukan adanya SK Menteri terkait untuk memberikan kepastian pasar sebagai pegangan bagi investor untuk mengembangkan industri bioetanol.
Posisi Penyuluh Pertanian Lapang (PPL) yang cukup solid pada saat revolusi hijau cenderung melemah dalam era otonomi daerah. Kondisi tersebut diperparah dengan beralih fungsinya PPL ke struktural dan PPL yang ada cenderung bias ke komoditas superior. Bila revitalisasi lembaga penyuluhan tidak dilaksanakan, keberhasilan pengembangan bahan bakar nabati untuk mengurangi ketergantungan terhadap BBM akan terhambat. Keputusan Presiden No. 05 dan Instruksi Presiden No. 1 tahun 2006 belum dilengkapi dengan SK tentang BBM mix, harga dan sistem distribusinya di SPBU belum merupakan kekuatan penuh bagi para investor untuk mengembangkan industri bioetanol. Bila Perda dan SK Menteri terkait tidak segera direalisasikan percepatan pengembangan industri bioetanol akan terhambat.
Tata ruang industri bioetanol yang ideal adalah yang berbasis daya dukung lingkungan yakni dalam bentuk hamparan lahan yang produksi ubi kayunya mampu memenuhi kebutuhan bahan baku harian selama setahun secara kontinyu. Sebagai contoh adalah setiap hamparan pertanaman ubi kayu dengan produktivitas 20 ton/ha dalam radius 30 km hanya boleh dibangun satu industri bioetanol dengan kapasitas 60 kl/hari. Dalam hamparan tersebut termasuk perkebunan energi yang dikelola oleh industri sebagai penyangga ketersediaan bahan baku. Ketentuan tersebut harus
termasuk dalam Perda. Koordinasi antar instansi terkait dengan progaram aksi pengembangan industri bioetanol dari pusat sampai daerah belum terwujud dalam suatu
kesepakatan. Bila hal tersebut tidak segera terimplementasi berpotensi menghambat investor untuk mengembangkan industri bioetanol di daerah sentra produksi ubi kayu.
Produktivitas ubi kayu yang mempunyai keunggulan dibanding komoditas tanaman pangan lain dalam usaha tani dipengaruhi oleh penggunaan teknologi inovatif spesifik lokasi. Teknologi produksi tersebut disusun berdasarkan hasil kajian dari BPTP bekerjasama dengan instansi terkait yang disusun oleh tim teknis. Otonomi daerah yang organisasinya inkonsisten mengakibatkan koordinasi tim teknis juga tidak konsisten. Bila hal tersebut tidak dibenahi berpotensi menghambat partisipasi petani dalam pengem- bangan industri bioetanol.
Petani produsen ubi kayu mengalami kesulitan dalam pemasaran hasil untuk mendapatkan harga yang layak, disebabkan oleh dukungan fasilitas dan infrastruktur yang belum memadai. Implikasinya adalah, industri bioetanol akan mengalami kesulitan untuk mendapatkan bahan baku dengan harga layak pada musim hujan disebabkan oleh sulitnya armada angkutan mencapai lahan petani yang infrastrukturnya belum memadai. Industri tapioka tidak tersebar sesuai dengan sebaran konsumen. Industri tapioka skala besar terkonsentrasi di Sumatera bagian selatan.
Peluang Pasar
konsumen tersebar di berbagai pulau. Kondisi tersebut berpotensi men-
dorong terjadinya harga tapioka impor dari Thailand lebih murah di daerah yang letaknya lebih dekat dengan Thailand dibanding Lampung. Hal tersebut berpeluang terjadi pada industri bioetanol bila sebaran industri bioetanol seperti sebaran industri tapioka, sehingga harga bioetanol akan bervariasi dan tidak kompetitif.
Strategi untuk mengatasi masalah manajemen tersebut diprioritaskan
pada:
• penegakan Perpres/Inpres agar lebih konsisten untuk mendorong minat
swasta mengembangkan industri bioetanol, dan melalui revitalisasi perta-
nian dan perhatian pemerintah yang besar untuk memenuhi permintaan
bioetanol yang tinggi,
• pemanfaatan upaya Pemda menegakkan Perpres No. 5 tahun 2006/Inpres
No. 1 tahun 2006 yang makin konsisten dan pengambilan keputusan melalui
Perda untuk meningkatkan koordinasi antar instansi terkait, dan permintaan
bioetanol yang tinggi untuk memperbaiki implementasi program pengem-
bangan ubi kayu sebagai bahan baku industri bioetanol,
• pemanfaatan perhatian pemerintah yang besar dan minat swasta
mengembangkan industri bioetanol untuk mencegah ekspor gaplek sebagai
bahan baku industri bioetanol skala besar di RRC, dan melalui revitalisasi
pertanian ekspor bioetanol oleh investor asing dan ekspor alkohol dapat
dicegah, dan
• revitalisasi lembaga penyuluhan dan perbaikan koordinasi antar instansi
yang terkait dengan pengembangan ubi kayu sebagai bahan baku industri
bioetanol untuk mencegah ekspor gaplek sebagai bahan baku industri
bioetanol RRC, ekspor bioetanol dan alkohol.
Aspek Biofisik
Hambatan yang terkait dengan aspek biofisik dalam pengembangan ubi kayu untuk memenuhi kebutuhan bahan baku industri bioetanol meliputi sumberdaya tanaman, lahan, dan petani. Permasalahan sumberdaya tanaman meliputi: (1) siklus pertanaman panjang, (2) pertumbuhan fase awal lambat, (3) kadar pati bervariasi dan spesifik agroekologi, (4) sifat hasil “bulky”. Permasalahan sumberdaya lahan meliputi: (1) kadar hara makro dan bahan organik tanah rendah, (2) peka erosi dan konservasinya belum efektif, (3) PTT belum berkembang, (4) sentra produksi didominasi oleh iklim kering, (5) kinerja kelompok tani lemah, (6) tenaga kerja produktif untuk usaha tani terbatas, dan (7) tingkat pendidikan rendah.
Dampak negatif dari umur tanaman yang panjang dalam usaha tani ubi kayu berdasarkan hasil pengamatan lapangan di daerah sentra produksi, diantaranya adalah: (1) investasi input rendah, (2) sering dipanen sebelum umur optimal, (3) sistem usaha tani subsisten, (4) didominasi oleh sistem tumpang sari untuk petani berlahan sempit, (5) sistem rotasi dengan tanaman pangan berumur pendek, dan (6) pengelolaan kurang optimal. Usaha tani ubi kayu dengan sistem tersebut produktivitasnya rendah dan cenderung tidak mempunyai keunggulan. Kondisi tersebut dapat menghambat pemenuhan bahan baku industri bioetanol. Pertumbuhan ubi kayu pada fase awal yang lambat berpotensi memicu munculnya masalah seperti erosi dan gangguan gulma. Erosi akan berlangsung sejalan dengan waktu yang menyebabkan terjadinya degradasi tanah bila tidak dilakukan konservasi yang efektif. Pertumbuhan ubi kayu yang lambat pada fase awal memacu gulma untuk tumbuh lebih cepat dan pertumbuhan ubi kayu terhambat karena tidak mampu berkompetisi dengan gulma. Bila pengendalian erosi dan gulma tidak dilakukan secara efektif, produktivitas ubi kayu tidak optimal sehingga berpotensi tidak terpenuhinya kebutuhan bahan baku untuk industri bioetanol.
Kadar pati maksimal berdasarkan potensi genetis dari varietas unggul dapat dicapai pada kondisi lingkungan optimal, dan akan menurun bila terjadi cekaman lingkungan. Kondisi optimal untuk tiap varietas tidak sama, sehingga penggunaan multivarietas unggul (umur genjah, sedang dan dalam) diperlukan pada daerah yang kondisi lingkungan abiotiknya bervariasi, seperti kemarau panjang atau anomali iklim, dan lain-lain.
Sistem tumpangsari ubi kayu dengan tanaman pangan lainnya berpotensi
menurunkan produktivitasnya bila terjadi kompetisi hara dan cahaya, namun berpeluang meningkatkan produktivitas bila pengaturan jenis tanaman dan populasi tanaman/jarak tanamnya tepat. Oleh karena jumlah petani berlahan sempit mendominasi petani di pulau Jawa, maka kondisi tersebut berpotensi mengganggu ketersediaan bahan baku industri bioetanol.
Sebagian lahan pertanian berkadar hara makro dan bahan organik antara rendah – sangat rendah. Hal tersebut dapat menjadi faktor pembatas produktivitas. Bila kondisi tersebut tidak diatasi, maka dapat menjadi faktor penghambat dalam upaya meningkatkan produksi ubi kayu untuk mendukung pengembangan industri bioetanol. Upaya tersebut sulit diimplementasikan disebabkan terbatasnya modal usaha tani dan petani yang memiliki ruminansia sebagai produsen pupuk organik jumlahnya relatif sedikit.Iklim kering mendominasi daerah sentra produksi ubi kayu. Karakteristik produksi ubi kayu di daerah beriklim kering adalah petaninya menanam ubi kayu secara serentak pada awal musim hujan dan bulan panen terkonsentrasi
pada musim kemarau. Dengan demikian produksi ubi kayu tidak terdistribusi merata sepanjang tahun sesuai dengan kebutuhan bahan baku industri bioetanol.Rataan hasil ubi kayu nasional sekitar 40–60% dari potensi genetis merupakan indikator bahwa pengelolaan tanaman terpadu (PTT) belum berkembang. Lahan marginal dan usaha tani subsisten yang mendominasi sebagian besar daerah sentra produksi ubi kayu merupakan penyebab utama belum berkembangnya teknologi pengelolaan tanaman terpadu/sistem integrasi ubi kayu–ternak (PTT/SIUT).
Kelembagaan Lokal dan SDM
Proyek Kemitraan Terpadu (PKT) adalah suatu program kemitraan terpadu yang melibatkanusaha besar (inti), usaha kecil (plasma) dengan melibatkan bank sebagai pemberi kreditdalam suatu ikatan kerja sama yang dituangkan dalam nota kesepakatan. Tujuan PKTantara lain adalah untuk meningkatkan kelayakan plasma, meningkatkan keterkaitan dankerjasama yang saling menguntungkan antara inti dan plasma, serta membantu bankdalam meningkatkan kredit usaha kecil secara lebih aman dan efisien.Dalam melakukan kemitraan hubunga kemitraan, perusahaan inti (Industri Pengolahan atauEksportir) dan petani plasma/usaha kecil mempunyai kedudukan hukum yang setara.Kemitraan dilaksanakan dengan disertai pembinaan oleh perusahaan inti, dimulai daripenyediaan sarana produksi, bimbingan teknis dan pemasaran hasil produksi.
Proyek Kemitraan Terpadu ini merupakan kerjasama kemitraan dalam bidang usahamelibatkan tiga unsur, yaitu (1) Petani/Kelompok Tani atau usaha kecil, (2) PengusahaBesar atau eksportir, dan (3) Bank pemberi KKPA.Masing-masing pihak memiliki peranan di dalam PKT yang sesuai dengan bidang usahanya.Hubungan kerjasama antara kelompok petani/usaha kecil dengan Pengusaha Pengolahanatau eksportir dalam PKT, dibuat seperti halnya hubungan antara Plasma dengan Inti didalam Pola Perusahaan Inti Rakyat (PIR). Petani/usaha kecil merupakan plasma danPerusahaan Pengelolaan/Eksportir sebagai Inti. Kerjasama kemitraan ini kemudian menjaditerpadu dengan keikut sertaan pihak bank yang memberi bantuan pinjaman bagipembiayaan usaha petani plasma. Proyek ini kemudian dikenal sebagai PKT yang disiapkandengan mendasarkan pada adanya saling berkepentingan diantara semua pihak yang bermitra.
1. Petani Plasma
Sesuai keperluan, petani yang dapat ikut dalam proyek ini bisa terdiri atas (a) Petani yangakan menggunakan lahan usaha pertaniannya untuk penanaman dan perkebunan atauusaha kecil lain, (b) Petani /usaha kecil yang telah memiliki usaha tetapi dalam keadaanyang perlu ditingkatkan dalam untuk itu memerlukan bantuan modal.Untuk kelompok (a), kegiatan proyek dimulai dari penyiapan lahan dan penanaman ataupenyiapan usaha, sedangkan untuk kelompok (b), kegiatan dimulai dari telah adanya kebunatau usaha yang berjalan, dalam batas masih bisa ditingkatkan produktivitasnya denganperbaikan pada aspek usaha.Luas lahan atau skala usaha bisa bervariasi sesuai luasan atau skala yang dimiliki olehmasing-masing petani/usaha kecil. Pada setiap kelompok tani/kelompok usaha, ditunjukseorang Ketua dan Sekretaris merangkap Bendahara. Tugas Ketua dan Sekretaris Kelompokadalah mengadakan koordinasi untuk pelaksanaan kegiatan yang harus dilakukan oleh parapetani anggotanya, didalam mengadakan hubungan dengan pihak Koperasi dan instansilainnya yang perlu, sesuai hasil kesepakatan anggota. Ketua kelompok wajib menyelenggarakan pertemuan kelompok secara rutin yang waktunya ditentukan berdasarkan kesepakatan kelompok.
2. Koperasi
Para petani/usaha kecil plasma sebagai peserta suatu PKT, sebaiknya menjadi anggotasuata koperasi primer di tempatnya. Koperasi bisa melakukan kegiatan-kegiatan untuk membantu plasma di dalam pembangunan kebun/usaha sesuai keperluannya. Fasilitas KKPA hanya bisa diperoleh melalui keanggotaan koperasi. Koperasi yang mengusahakanKKPA harus sudah berbadan hukum dan memiliki kemampuan serta fasilitas yang cukupbaik untuk keperluan pengelolaan administrasi pinjaman KKPA para anggotanya. Jikamenggunakan skim Kredit Usaha Kecil (KUK), kehadiran koperasi primer tidak merupakankeharusan
3. Perusahaan Besar dan Pengelola/Eksportir
Suatu Perusahaan dan Pengelola/Eksportir yang bersedia menjalin kerjasama sebagai intidalam Proyek Kemitraan terpadu ini, harus memiliki kemampuan dan fasilitas pengolahanuntuk bisa menlakukan ekspor, serta bersedia membeli seluruh produksi dari plasma untukselanjutnya diolah di pabrik dan atau diekspor. Disamping ini, perusahaan inti perlumemberikan bimbingan teknis usaha dan membantu dalam pengadaan sarana produksiuntuk keperluan petani plasma/usaha kecil.Apabila Perusahaan Mitra tidak memiliki kemampuan cukup untuk mengadakan pembinaanteknis usaha, PKT tetap akan bisa dikembangkan dengan sekurang-kurangnya pihak Intimemiliki fasilitas pengolahan untuk diekspor, hal ini penting untuk memastikan adanyapemasaran bagi produksi petani atau plasma. Meskipun demikian petani plasma/usaha kecildimungkinkan untuk mengolah hasil panennya, yang kemudian harus dijual kepadaPerusahaan Inti.Dalam hal perusahaan inti tidak bisa melakukan pembinaan teknis, kegiatan pembibinganharus dapat diadakan oleh Koperasi dengan memanfaatkan bantuan tenaga pihak DinasPerkebunan atau lainnya yang dikoordinasikan oleh Koperasi. Apabila koperasimenggunakan tenaga Penyuluh Pertanian Lapangan (PPL), perlu mendapatkan persetujuanDinas Perkebunan setempat dan koperasi memberikan bantuan biaya yang diperlukan.Koperasi juga bisa memperkerjakan langsung tenaga-tenaga teknis yang memilikiketerampilan dibidang perkebunan/usaha untuk membimbing petani/usaha kecil dengandibiayai sendiri oleh Koperasi. Tenaga-tenaga ini bisa diberi honorarium oleh Koperasi yangbisa kemudian dibebankan kepada petani, dari hasil penjualan secara proposional menurutbesarnya produksi. Sehingga makin tinggi produksi kebun petani/usaha kecil, akan semakinbesar pula honor yang diterimanya.
4. Bank
Bank berdasarkan adanya kelayakan usaha dalam kemitraan antara pihak Petani Plasmadengan Perusahaan Perkebunan dan Pengolahan/Eksportir sebagai inti, dapat kemudianmelibatkan diri untuk biaya investasi dan modal kerja pembangunan atau perbaikan kebun.Disamping mengadakan pengamatan terhadap kelayakan aspek-aspek budidaya/produksiyang diperlukan, termasuk kelayakan keuangan. Pihak bank di dalam mengadakan evaluasi, juga harus memastikan bagaimana pengelolaan kredit dan persyaratan lainnya yang diperlukan sehingga dapat menunjang keberhasilan proyek. Skim kredit yang akandigunakan untuk pembiayaan ini, bisa dipilih berdasarkan besarnya tingkat bunga yangsesuai dengan bentuk usaha tani ini, sehingga mengarah pada perolehannya pendapatanbersih petani yang paling besar.Dalam pelaksanaanya, Bank harus dapat mengatur cara petani plasma akan mencairkankredit dan mempergunakannya untuk keperluan operasional lapangan, dan bagaimanapetani akan membayar angsuran pengembalian pokok pinjaman beserta bunganya. Untukini, bank agar membuat perjanjian kerjasama dengan pihak perusahaan inti, berdasarkankesepakatan pihak petani/kelompok tani/koperasi. Perusahaan inti akan memotong uanghasil penjualan petani plasma/usaha kecil sejumlah yang disepakati bersama untukdibayarkan langsung kepada bank. Besarnya potongan disesuaikan dengan rencanaangsuran yang telah dibuat pada waktu perjanjian kredit dibuat oleh pihak petani/Kelompoktani/koperasi. Perusahaan inti akan memotong uang hasil penjualan petani plasma/usahakecil sejumlah yang disepakati bersama untuk dibayarkan langsung kepada Bank. Besarnyapotongan disesuaikan dengan rencana angsuran yang telah dibuat pada waktu perjanjiankredit dibuat oleh pihak petani plasma dengan bank.
Pola Kerjasama
Kemitraan antara petani/kelompok tani/koperasi dengan perusahaan mitra, dapat dibuatmenurut dua pola yaitu :a. Petani yang tergabung dalam kelompok-kelompok tani mengadakan perjanjian kerjasamalangsung kepada Perusahaan Perkebunan/Pengolahan Eksportir.Dengan bentuk kerja sama seperti ini, pemberian kredit yang berupa KKPA kepada petaniplasma dilakukan dengan kedudukan Koperasi sebagai Channeling Agent, danpengelolaannya langsung ditangani oleh Kelompok tani. Sedangkan masalah pembinaanharus bisa diberikan oleh Perusahaan Mitra.b. Petani yang tergabung dalam kelompok-kelompok tani, melalui koperasinya mengadakanperjanjian yang dibuat antara Koperasi (mewakili anggotanya) dengan perusahaanperkebunan/pengolahan/eksportir.
Petani yang tergabung dalam kelompok-kelompok tani mengadakan perjanjian kerjasamalangsung kepada Perusahaan Perkebunan/Pengolahan Eksportir.Dengan bentuk kerja sama seperti ini, pemberian kredit yang berupa KKPA kepada petaniplasma dilakukan dengan kedudukan Koperasi sebagai Channeling Agent, danpengelolaannya langsung ditangani oleh Kelompok tani. Sedangkan masalah pembinaanharus bisa diberikan oleh Perusahaan Mitra.b. Petani yang tergabung dalam kelompok - kelompok tani, melalui koperasinya mengadakan perjanjian yang dibuat antara Koperasi (mewakili anggotanya) dengan perusahaanperkebunan/pengolahan/eksportir.
Dalam bentuk kerjasama seperti ini, pemberian KKPA kepada petani plasma dilakukandengan kedudukan koperasi sebagai Executing Agent. Masalah pembinaan teknis budidayatanaman/pengelolaan usaha, apabila tidak dapat dilaksanakan oleh pihak Perusahaan Mitra,akan menjadi tanggung jawab koperasi.
Penyiapan Proyek Kemitraan Terpadu
Untuk melihat bahwa PKT ini dikembangkan dengan sebaiknya dan dalam proseskegiatannya nanti memperoleh kelancaran dan keberhasilan, minimal dapat dilihat daribagaimana PKT ini disiapkan. Kalau PKT ini akan mempergunakan KKPA untuk modal usahaplasma, perintisannya dimulai dari
A. Adanya petani/pengusaha kecil yang telah menjadi anggota koperasi dan lahan pemilikannya akan dijadikan kebun/tempat usaha atau lahan kebun/usahanya sudah adatetapi akan ditingkatkan produktivitasnya. Petani/usaha kecil tersebut harus menghimpundiri dalam kelompok dengan anggota sekitar 25 petani/kelompok usaha. Berdasarkan persetujuan bersama, yang didapatkan melalui pertemuan anggota kelompok, merekabersedia atau berkeinginan untuk bekerja sama dengan perusahaanperkebunan/pengolahan/eksportir dan bersedia mengajukan permohonan kredit (KKPA)untuk keperluan peningkatan usaha. Adanya perusahaan perkebunan/pengolahan dan eksportir, yang bersedia menjadimitra petani/usaha kecil, dan dapat membantu memberikan pembinaan teknikbudidaya/produksi serta proses pemasarannya, Dipertemukannya kelompok tani/usaha kecil dan pengusaha perkebunan/pengolahandan eksportir tersebut, untuk memperoleh kesepakatan di antara keduanya untuk bermitra.Prakarsa bisa dimulai dari salah satu pihak untuk mengadakan pendekatan, atau ada pihakyang akan membantu sebagai mediator, peran konsultan bisa dimanfaatkan untukmengadakan identifikasi dan menghubungkan pihak kelompok tani/usaha kecil yangpotensial dengan perusahaan yang dipilih memiliki kemampuan tinggi memberikan fasilitasyang diperlukan oleh pihak petani/usaha kecil;
Diperoleh dukungan untuk kemitraan yang melibatkan para anggotanya oleh pihakkoperasi. Koperasi harus memiliki kemampuan di dalam mengorganisasikan dan mengelolaadministrasi yang berkaitan dengan PKT ini. Apabila keterampilan koperasi kurang, untukpeningkatannya dapat diharapkan nantinya mendapat pembinaan dari perusahaan mitra.Koperasi kemudian mengadakan langkah-langkah yang berkaitan dengan formalitas PKTsesuai fungsinya. Dalam kaitannya dengan penggunaan KKPA, Koperasi harus mendapatkanpersetujuan dari para anggotanya, apakah akan beritndak sebagai badan pelaksana(executing agent) atau badan penyalur (channeling agent);
Diperolehnya rekomendasi tentang pengembangan PKT ini oleh pihak instansipemerintah setempat yang berkaitan (Dinas Perkebunan, Dinas Koperasi, KantorBadan Pertanahan, dan Pemda);
Mekanisme Proyek Kemitraan Terpadu
Mekanisme Proyek Kemitraan Terpadu dapat dilihat pada skema berikut ini Bank pelaksana akan menilai kelayakan usaha sesuai dengan prinsip-prinsip bank teknis.Jika proyek layak untuk dikembangkan, perlu dibuat suatu nota kesepakatan (Memorandumof Understanding = MoU) yang mengikat hak dan kewajiban masing-masing pihak yang bermitra (inti, Plasma/Koperasi dan Bank). Sesuai dengan nota kesepakatan, atas kuasakoperasi atau plasma, kredit perbankan dapat dialihkan dari rekening koperasi/plasma kerekening inti untuk selanjutnya disalurkan ke plasma dalam bentuk sarana produksi, danapekerjaan fisik, dan lain-lain.
Dengan demikian plasma tidak akan menerima uang tunai dariperbankan, tetapi yang diterima adalah sarana produksi pertanian yang penyalurannyadapat melalui inti atau koperasi. Petani plasma melaksanakan proses produksi. Hasiltanaman plasma dijual ke inti dengan harga yang telah disepakati dalam MoU. Perusahaaninti akan memotong sebagian hasil penjualan plasma untuk diserahkan kepada banksebagai angsuran pinjaman dan sisanya dikembalikan ke petani sebagai pendapatan bersih.
DAFTAR PUSTAKA
Anonim, 2000. Ketika Kebun Berupa Hutan. Agroforestri Khas Indonesia.
Sebuah Sumbangan Masyarakat. International Centre For Research In
Agroforestry. Bogor.
Anonim. 2001. Sistem Pertanian di Indonesia.http://www.lab link.or.id. Diakses
pada tanggal 22 Mei 2010 pukul 17.00 WIB
Danoesastro, Haryono. 1979. Pemanfaatan Pekarangan. Yayaan Pembina FakulasPertanian UGM. Yogyakarta
.
Dover,M. dan Talbot,L.M., 1987. To Feed The Earth: Agroecology for
Sustainable Development. World Resources Intitute. Washington DC.
Handayanto, E. 1999. Pengelolan Kesuburan Tanah. Fakultas Pertanian.
Universitas Brawijaya. Malang.
Hardjowigeno, S., 1989. Ilmu Tanah. Mediyatama Sarana Perkasa, Jakarta
Manuwoto.2009. Sistem Pertanian di Indonesia.
Http://makhey.blogspot.com/2009/09/sistem-pertanian-di-indonesia.
Diakses pada tanggal 27 Mei 2010.
Monika, WT et al. 1993. Produksi Kambing dan Domba di Indonesia. Sebelas
Maret Universitas Press. Surakarta.
Notohadinagoro, Tejoyuwono. 1997. Bercari manat Pengelolaan Berkelanjutan
Sebagai Konsep Pengembangan Wilayah Lahan Kering.Makalah
Seminar Nasional dan Pelatihan Pengelolaan Lahan Kering FOKUSHIMITI di Jember. Universitas Jember. Jember
Pusat Peneliti Universitas Brawijaya. 1991. Penelitian dan Pengembangan Sistem
Usaha Tani Lahan Kering Yang Berkelanjutan. Proseding Simposium Nasional Malang. Universitas Brawijaya. Malang
Reijntjes,C., B.Haverkot dan A. W. Bayer., 1999. Pertanian Masa Depan Pengantar untuk Pertanian Berkelanjutan Dengan Input Luar Rendah. Kanisius. Yogyakarta.
TUGAS KELOMPOK
SISTEM PERTANIAN TERPADU
EKOSISTEM TANAMAN UBI KAYU
DISUSUN OLEH :
ERNAWATY DJAYA ( G111 09 276)
RATI PRADHANI ( G111 09 294 )
NUR JANNAH
IRAWAN TAMRIN
PROGRAM STUDI AGROTEKNOLOGI
JURUSAN ILMU HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
UNIVERSITAS HASANUDDIN
MAKASSAR
2011
Langganan:
Postingan (Atom)